

Une nouvelle approche de la fertilisation des cultures
Gilles Lemaire
Ce texte a été publié dans sa version originale dans Les mots de l’agronomie, et peut être téléchargé sur le lien https://mots-agronomie.inra.fr/index.php/Fertilisation_des_cultures_:_des_bases_scientifiques_renouvel%C3%A9es
Résumé
Les pratiques de fertilisation en agriculture basées sur une approche de pronostic de la réponse des cultures à l’apport d’éléments fertilisants ont permis les augmentations considérables des rendements depuis plus d’un siècle. La forte incertitude attachée à ces pronostics, même réduite autant que possible à l’aide d’un diagnostic basé sur les analyses de sol, a conduit les prescripteurs et les agriculteurs à maximiser les doses d’apport d’engrais du fait de leur aversion au risque d’une sous-fertilisation des cultures. Cette pratique généralisée a été la cause d’une mise en circulation excessive d’azote et de phosphore dans les agro-écosystèmes qui est responsable d’impacts environnementaux négatifs importants qui ne sont plus acceptés par la société.
Un changement de paradigme, considérant la disponibilité des éléments nutritifs pour la plante, non plus comme un facteur externe à celle-ci, uniquement contrôlé par les apports de fertilisants et par leurs interactions physico-chimiques avec la matrice minérale du sol, mais comme une propriété émergente du système plante – sol – microbiome* [1] , permet de mettre en œuvre un approche basée sur le diagnostic in situ de l’état de nutrition de la culture qui de fait intègre la variabilité contextuelle et permet donc d’inférer avec une incertitude beaucoup plus faible les besoins en apports d’engrais des cultures et du même coup les risques environnementaux qui leur sont associés.
Introduction
Depuis le début du 20e siècle, les rendements en grain des principales cultures nécessaires directement ou indirectement à l’alimentation humaine ont été multipliés par un facteur 10, en grande partie grâce à une augmentation d’un facteur 7 de l’utilisation des engrais azotés et d’un facteur 3,5 de celle des engrais phosphatés (Tilman et al., 2002). Il s’agit donc d’un indéniable succès permettant de nourrir une population humaine toujours croissante sur des surfaces cultivables limitées.
Cependant ce succès atteint aujourd’hui ses limites pour au moins deux raisons majeures :
(i) La limitation des ressources en éléments fertilisants du fait que l’azote (N), essentiellement fourni par le procédé Haber-Bosch à partir d’un réservoir inépuisable, l’azote atmosphérique (N2), est en réalité une ressource limitée à l’avenir du fait de son coût en énergie fossile et des émissions de gaz à effet de serre qui accompagnent ce procédé industriel ; et du fait que le phosphore (P) est extrait de minerais dont la ressource est forcément limitée à l’échelle du globe.
(ii) La faible efficience d’utilisation de ces deux éléments dans les systèmes agricoles, qui conduit à des augmentations excessives de leur circulation sous forme réactive au sein des agro-écosystèmes provoquant des impacts négatifs en cascade dans l’environnement (Galloway & Cowling, 2002).
Les recherches sur la nutrition minérale et la fertilisation des cultures ont été menées sur la base des paradigmes* issus des travaux pionniers de Boussingault (1855) et de Liebig (1855) au milieu du XIXe siècle. Ces paradigmes avaient pour objet la réponse des plantes et des cultures à des apports exogènes d’éléments minéraux sous formes d’engrais. L’aspect endogène de la disponibilité de ces éléments dans la dynamique de fonctionnement du système sol – plante – microbes n’était donc pas pris en compte, et par conséquent les courbes de réponse obtenues expérimentalement étaient fortement dépendantes du contexte local sol – climat - plante et donc éminemment variables dans l’espace et dans le temps. Face à cette variabilité, l’utilisation des courbes de réponse comme outils de pronostic pour l’établissement des règles de fertilisation a donc engendré une forte incertitude qui a incité les prescripteurs et les agriculteurs à minimiser les risques de sous-fertilisation en apportant des quantités d’engrais souvent excédentaires pour être certain de satisfaire « à coup sûr » les besoins de leurs cultures (Ravier et al., 2016). Cette pratique, encouragée par un faible prix des engrais comparé à ceux des produits agricoles, a conduit à des excès qui, cumulés au cours des ans, sont responsables des flux excessifs de N et P dans l’environnement observés aujourd’hui.
Il convient donc de re-questionner les paradigmes initiaux sur lesquels l’agronomie s’est en partie fondée en matière de nutrition minérale et de fertilisation des cultures, afin de réintroduire une vision plus dynamique du fonctionnement du système sol – plante - micro-organismes, qui puisse prendre en compte non seulement la réponse des plantes à des apports exogènes d’éléments nutritifs, mais aussi les nombreuses rétroactions endogènes qui confèrent au système ses propriétés adaptatives.
Les limites inhérentes à la Loi de Liebig, dite loi du minimum.
La « loi du minimum » de Liebig (1855) s’énonce de la manière suivante « Les plantes ne croissent qu’en fonction du niveau permis par l’élément nutritif qui est le plus limitant ». Son interprétation comme « loi des facteurs limitants » et sa traduction et son extension à l’ensemble des facteurs du milieu par les courbes dites de Blackman (1905) a fait naître l’idée que la croissance des plantes était contrainte par des facteurs limitants successifs, indépendants entre eux, agissant ou se révélant de manière séquentielle au fur et à mesure où l’on agit sur le milieu par des mesures correctives (Figure 1).
ERROR: Content Element with uid "307" and type "JBdesign_IMGalone" has no rendering definition!
Cette vision, complétée par la Loi de Mistcherlich (1924) qui montre que l’intensité de la réponse des plantes à un facteur donné est décroissante et donc asymptotique, a prévalu comme paradigme de base en agronomie et a été le cadre dans lequel s’est développé l’ensemble des recherches sur la fertilisation des cultures (Figure 2).
ERROR: Content Element with uid "305" and type "JBdesign_IMGalone" has no rendering definition!
Cependant, dès la fin du XIXe siècle, Liebscher (1885) avait proposé une formulation plus complète de ce que voulait signifier Liebig sous la forme de la loi dite de l’optimum : « Chaque élément nutritif est utilisé de manière d’autant plus efficiente que la disponibilité de l’élément qui est le plus limitant est portée près de son optimum », ce qui ouvrait la voie à une analyse plus globale en explicitant l’existence d’interactions entre les éléments, au-delà du simple effet « tout ou rien » de la loi de Liebig telle qu’elle avait été formulée et interprétée. Mais, curieusement, les travaux de Liebscher sont restés relativement méconnus, et c’est sur la base de la Loi de Liebig et son interprétation assez simplifiée que l’agronomie a été enseignée à travers le monde. Ceci a eu pour effet une division excessive des recherches sur la nutrition des plantes et la fertilisation des cultures en compartimentant la réponse des plantes à chacun des principaux éléments, N, P, K…micro- et oligo-éléments, allant même jusqu’à la spécialisation des équipes de recherches pour chaque élément particulier.
ERROR: Content Element with uid "303" and type "JBdesign_IMGalone" has no rendering definition!
L’analyse des courbes de réponse des cultures aux apports croissants d’éléments nutritifs à l’aide des outils statistiques et de l’expérimentation « au champ » a donc été la base des recherches en agronomie de 1950 à 1990 environ, ne traitant éventuellement des interactions entre éléments que par le biais des résidus de variations expérimentales non expliquées par les effets élémentaires.
Il est maintenant bien établi que la loi de Liebig ne permet pas de rendre compte des interactions entre tous les facteurs du milieu, non seulement les éléments nutritifs entre eux, mais aussi avec les autres ressources telles que l’eau (Sinclair & Park, 1993 ; Cossani & Sadras, 2018). La raison en est que la croissance des plantes ne résulte pas d’une séquence linéaire de processus, chacun d’eux étant limité par un facteur donné, mais plutôt de boucles de processus eux-mêmes gouvernés par plusieurs facteurs à la fois (Rabinovitch, 1951). Les recherches sur la nutrition des cultures en agronomie ont été tout naturellement dominées par cette vision linéaire et séquentielle des facteurs du milieu par le fait qu’elles étaient « finalisées et guidées » par l’étude de la réponse des cultures à une action anthropique comme la fertilisation. Dans le même temps et parallèlement, les recherches en Écologie fonctionnelle*, guidées par la seule compréhension du fonctionnement des couverts végétaux in natura, ont développé des approches très différentes basées sur le concept de co-limitation entre facteurs du milieu (Bloom et al., 1985 ; Kho, 2000). Ce concept de co-limitation se rapproche davantage de la vision de Liebscher que de celle de Liebig et permet d’expliciter formellement les nombreuses interactions et rétroactions qui entrent en jeu dans le fonctionnement du système plante-sol. Cette approche de modélisation dynamique du fonctionnement des couverts végétaux, initiée par les travaux de De Wit (1992, 1994) a permis d’expliciter les processus d’acquisition et d’allocation des ressources, ainsi que les concepts d’efficience de conversion des ressources qui sont à la base de tous les modèles actuels de fonctionnement des couverts végétaux et donc des cultures (Hanks & Ritchie, 1991 ; Monteith et al., 1994). Cette approche a été appliquée essentiellement aux ressources telles que la lumière, le CO2 ou l’eau, mais plus rarement aux éléments nutritifs.
La grande variabilité des courbes de réponses des cultures aux apports d’éléments nutritifs, obtenues expérimentalement par les agronomes a incité ceux-ci à spécifier les variables locales du sol susceptibles de résorber une partie de l’incertitude du « pronostic » qui pouvait en être déduit. La Science du Sol s’est donc tout naturellement emparée du problème des interactions entre les différents éléments nutritifs et la matrice minérale du sol, pour tenter de quantifier la « disponibilité » in situ de ces éléments pour les plantes. Ainsi l’approche physico-chimique de la nutrition minérale a engendré une interprétation plus mécaniste des courbes de réponse en proposant un « diagnostic du sol » comme co-variable contextuelle permettant d’inférer un « pronostic de nutrition » de la culture. Mais, restant dans le cadre conceptuel de la loi de Liebig, cette approche explicative physico-chimique a concentré son effort sur le concept de « disponibilité » de chacun des éléments pris séparément, alors que les concepts de co-limitation et de stoechiométrie* développés en Écologie fonctionnelle, qui permettaient une vision systémique de la nutrition des plantes, n’ont que très rarement été pris en compte par les agronomes (Reich et al., 2018).
Pour les éléments nutritifs les moins mobiles, ayant les plus fortes interactions avec la matrice minérale des sols, le concept de « disponibilité » a été formulé au moyen de valeurs seuils issues de tests physico-chimiques réalisés par des analyses de sol in vitro, essayant de traduire ainsi la capacité qu’auraient les plantes à prélever chacun des éléments indépendamment les uns des autres en fonction des interactions plus ou moins fortes de ces éléments avec la matrice minérale du sol. Cette approche normative, basée sur des valeurs seuils déterminées empiriquement, a permis un raisonnement des pratiques de fertilisation en fonction des types de sols et de leur histoire culturale, et a permis et accompagné la formidable expansion de la fertilisation des cultures et l’augmentation des rendements dans les agricultures développées. Il s’agit donc d’un indéniable succès. Cependant, le bilan de cette approche en termes de capacité prédictive pour la détermination de la dose optimale d’apport de fertilisant pour les différentes cultures reste encore aujourd’hui relativement limité (Valkama et al., 2011). Le principal écueil de cette approche normative est qu’elle considère la « disponibilité » de chacun des éléments séparément : elle ignore les synergies ou antagonismes entre éléments, mais aussi et surtout elle ne considère absolument pas les autres composantes du système, la plante elle-même et le compartiment biologique du sol, comme acteurs effectifs de cette « disponibilité ». En externalisant la disponibilité des éléments nutritifs, c’est-à-dire en la considérant comme une donnée du milieu « externe à la plante », cette approche ne peut en aucun cas prendre en compte les nombreux processus, interactions et rétroactions qui in fine déterminent la capacité qu’ont les plantes à absorber les éléments présents dans leur environnement racinaire. La disponibilité des éléments nutritifs doit au contraire être considérée comme résultante de la dynamique interne du système et comme l’expression de ses propriétés émergentes*.
Pour les éléments mobiles comme l’azote (N), les agronomes se sont très tôt aperçus qu’il convenait de faire coïncider les apports de fertilisants avec la dynamique du besoin des cultures, la matrice minérale du sol jouant alors un rôle très mineur. Là encore, l’approche linéaire entre apports sous forme d’engrais et réponses en terme de rendement a montré ses limites face à l’énorme variabilité des fournitures d’azote par la minéralisation des matières organiques du sol, et la non moindre variabilité des besoins instantanée des plantes (Bramley et al., 2013), ainsi que cela est illustré Figure 4.
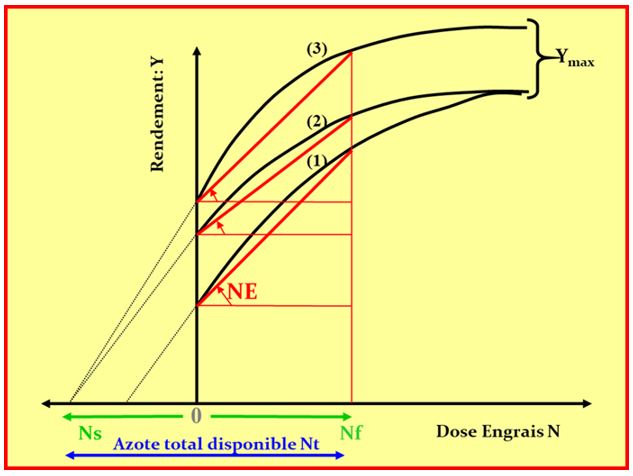
Figure 4 : Représentation des courbes de réponses du rendement (Y) d’une culture à des apports d’engrais azotée. La dose de N optimale (Nf) pour obtenir les rendements maxima Ymax dépend 1° de la fourniture de N par le sol (Ns) et 2° de la valeur de Ymax dépendant du climat et du génotype. L’ensemble de ces incertitudes engendrent une forte variation de l’efficience de l’azote (NE).
Afin de surmonter cet obstacle d’imprévisibilité des réponses des cultures à l’apport d’engrais azoté, les agronomes français ont tenté de prévoir la disponibilité en azote dans le sol pour les cultures en établissant un bilan prévisionnel de N minéral dans un contexte de sol et de climat donné (Hébert, 1969 ; Rémy & Hébert, 1977 ; Meynard et al., 1997). Cette approche, synthétisée par Machet et al. (2017), permet de prévoir approximativement la quantité d’azote qui serait « disponible » pour une culture donnée sur l’ensemble de son cycle de croissance, en faisant un bilan entre (i) un pronostic de la fourniture de N par la minéralisation de la matière organique du sol, (ii) une mesure de reliquat de N minéral dans le sol, et (iii) une estimation des pertes par volatilisation, dénitrification et lixiviation. Bien que cette méthode soit capable de réduire fortement l’incertitude de la réponse de la culture aux apports d’azote, sa capacité de prédiction dans une situation donnée reste trop imprécise et donne donc encore lieu à des pratiques de fertilisation de type « assurance » liées à l’aversion aux risques des agriculteurs (Ravier et al., 2016 ; 2018), conduisant à des fertilisations excédentaires qui restent encore non compatibles avec la préservation de l’environnement (Meynard et al., 2002).
Aujourd’hui, les approches de type « pronostics », qu’elles soient basées sur des tests de type « analyses de sols » pour P, K et autres minéraux, ou sur des modélisations empiriques de bilans de N, conduisent à des décisions en matière de fertilisation des cultures qui sont en règle général excédentaires du seul fait de la grande incertitude qui reste attachée à la prévision dans un contexte particulier. Il en découle donc une incapacité à ajuster les apports de fertilisants au strict besoin des cultures, causant des impacts environnementaux importants dans les régions d’agriculture intensive, alors que d’autres régions du monde restent largement sous-fertilisées (Ringeval et al., 2017).
Pour une approche du diagnostic in situ de l’état de nutrition des cultures
Si les pronostics basés sur des « diagnostics sol », que ce soit de type analyses de sols P, K etc… ou de type « bilan N », s’avèrent trop imprécis pour fonder des décisions en matière de fertilisation des cultures, pourquoi alors ne pas interroger directement les plantes in situ pour en déduire leur état de nutrition minérale ? C’est ce que permet théoriquement le concept de concentration critique des plantes en élément nutritif défini par Ulrich (1952) comme « le minimum de concentration en un élément minéral particulier nécessaire pour obtenir la croissance maximale de la plante ». Donc, en comparant à chaque instant la concentration réelle de la plante en un minéral particulier (% M) à la concentration critique (% Mc), il doit être possible de qualifier et de quantifier une carence ou un excès de nutrition en cet élément. C’est le principe du diagnostic foliaire (Smith, 1962 ; Cornforth et al., 1981) qui a été particulièrement mis en œuvre aux USA (Walworth & Summer, 1987), permettant notamment un diagnostic multi-éléments des cultures in situ. C’est aussi le principe du diagnostic azoté des cultures utilisant les propriétés de réflectance des couverts végétaux qui peuvent être corrélées à la concentration en N des plantes (Barret & Fourty, 1994 ; Cao et al., 2016), visant un pilotage en temps réel des apports d’azote grâce à des mesures de télédétection*.
Mais ces diagnostics « plante » ont longtemps buté sur l’absence de références stables et suffisamment génériques en termes de concentration critique % Mc. Ces valeurs critiques se sont avérées assez variables en fonction des espèces et cultivars, des stades phénologiques, des saisons, etc., donnant lieu à des tentatives nombreuses et infructueuses de normalisation et de définition de valeurs seuils, qui ont rendu cette approche rapidement très décevante malgré sa simplicité apparente.
Pendant ce temps, les recherches en écologie fonctionnelle sur les végétations naturelles forestières ou prairiales ont fait ressortir les aspects intégratifs et adaptatifs de l’acquisition, de l’allocation et du recyclage des éléments nutritifs au sein de l’écosystème (Blum, 1977 ; Agren, 2004 ; Niklas et al., 2005). Bien que rarement utilisée par les agronomes, l’allométrie*, c’est-à-dire l’analyse de croissance comparée des divers compartiments d’un organisme vivant, a permis de rendre compte des rétro-contrôles qui lient l’acquisition et l’allocation des ressources à la capacité de croissance des plantes. Ainsi les études de Lemaire et Salette (1984) sur les peuplements de graminées fourragères, de Lemaire et al. (1985) sur la luzerne et leur extension par Greenwood et al. (1990) à un large panel d’espèces en C3 et C4 cultivées ont débouché sur le concept de courbes de dilution de l’azote (Lemaire et al., 2008), montrant que la teneur en N critique des plantes diminue avec l’accumulation de biomasse du peuplement végétal du fait d’une allométrie entre prélèvement d’azote et accumulation de biomasse. Ainsi une interprétation de la concentration en azote des plantes à un moment donné dans une culture donnée (% Nact) peut être faite en référence à une valeur critique (% Ncrit) grâce à un indice de nutrition azotée INN = % Nact / % Ncrit, facilement mesurable à tout instant et dont une valeur inférieure à 1 indique et quantifie le déficit de nutrition azotée de la culture, alors qu’une valeur supérieure à 1 quantifie un excès ou une « consommation de luxe » d’azote. (Figure 5)
ERROR: Content Element with uid "299" and type "JBdesign_IMGalone" has no rendering definition!
Initialement développée de manière empirique, cette approche a été progressivement étayée par une analyse théorique de la nutrition azotée des plantes, qui met en exergue un certain nombre de régulations internes au système plante-sol :
(i) Une co-régulation de l’absorption d’azote par la concentration en N minéral du milieu racinaire et par la vitesse de croissance potentielle de la plante elle-même, qui établit que la « disponibilité en azote du sol » dépend aussi de la plante elle-même (Lejay et al., 1999 ; Devienne-Barret, 2000).
(ii) Une accumulation d’azote dans la plante, qui est proportionnelle à la taille de son compartiment métabolique, lequel est déterminé par sa surface foliaire éclairée (Caloin & Yu, 1984 ; Lemaire et al,. 2007).
(iii) Une allocation d’azote dans la plante qui est sous la dépendance du développement architectural de la plante, sans lien direct avec des stades phénologiques (Ratgen et al., 2018).
De cet ensemble de données écophysiologiques, il ressort deux conclusions importantes :
(i) La disponibilité en azote ne peut plus être considérée comme un facteur externe aux variations duquel la plante (et donc la culture) exprimerait une réponse, mais comme une résultante interne du fonctionnement du système sol - plante, puisque la plante elle-même détermine sa capacité de prélèvement d’azote par sa propre vitesse de croissance, déterminée à la fois par son génotype et par le climat instantané auquel elle est soumise. Donc, d’une approche typiquement linéaire de type « action-réponse », on doit passer à une approche circulaire et dynamique où les effets rétroagissent constamment sur les causes.
(ii) Le diagnostic nutritionnel de la plante ne peut plus se faire de manière univoque par la seule détermination de sa concentration en azote puisque celle-ci évolue avec la croissance de la plante. Il convient donc de comparer des cultures de même biomasse et donc à même niveau de dilution de l’azote.
Une revue récente (Lemaire et al., 2019) montre l’emboîtement théorique étroit entre les échelles de la plante et du peuplement, expliquant la généricité des relations d’allométrie entre la dynamique de croissance d’une culture et sa dynamique de prélèvement d’azote et explicitant du même coup les propriétés émergentes observables à l’échelle de la culture. Cette même revue montre également la transposition du concept de dilution tel qu’il a été conçu pour l’azote, aux autres éléments minéraux impliqués dans le métabolisme de la plante, P, K et S. Il est ainsi possible de déterminer, à l’instar de l’INN pour l’azote, des indices de nutrition P (INP) ou K (INK) à partir de la définition des courbes critiques de dilution P et K (Duru & Thellier, 1997 ; Salette e& Huché, 1991) et aussi de rendre compte des interactions N-P et N-K par la détermination d’indices P/N ou K/N montrant que ces éléments sont en étroite interaction, la disponibilité de l’un ayant une influence importante sur la disponibilité de l’autre. (Ziadi et al., 2007 & 2008).
Cette nouvelle vision de la nutrition minérale des cultures considère que les rétroactions entre processus au sein du système sol-plante-microbiome du sol se traduisent par des propriétés émergentes du système lui-même, ce qui lui confère des propriétés auto-adaptatives et évolutives qui ne peuvent s’analyser que de manière dynamique. Une telle approche doit donc remplacer la vision linéaire considérant la réponse du système à une sollicitation exercée sur un facteur externe à celui-ci, telle qu’elle a été conçue par les paradigmes initiaux de l’agronomie. La complexité de la dynamique de ce système, examinée des points de vue physico-chimique et biologique, en rend toute prévision d’évolution entachée d’une très grande incertitude. L’abandon d’une approche de « pronostic » et son remplacement par une approche de « diagnostic » apparaît donc nécessaire. Ceci est d’autant plus clair que les recherches récentes, tant en physiologie végétale qu’en écologie du sol, montrent que :
(1) Les plantes sont capables de percevoir directement des signaux concernant la présence et la concentration des éléments minéraux dans leur milieu, de transmettre des signaux de déficience ou de satiété en différents éléments nutritifs depuis leurs parties aériennes vers leurs racines et moduler ainsi en conséquence leur propre capacité d’absorption dans le cadre de réponses intégrées (Briat et al., 2015 & 2020 ; O’Brien et al., 2016; Ohkubo, 2017), rendant donc celles-ci actrices de la disponibilité de ces éléments dans le milieu;
(2) Le microbiome des sols, lui-même sous la dépendance nutritionnelle de la plante, rétroagit sur la biodisponibilité des éléments minéraux dans le sol (Briat et al., 2020), soit en l’augmentant grâce à ses capacités de captation dans le cadre de symbioses, soit au contraire comme compétiteurs vis-à-vis de la plante elle-même (immobilisation).
Ainsi, en ignorant la plante et le microbiome du sol comme agents effectifs de la disponibilité des éléments nutritifs dans le sol, les paradigmes qui ont fondé les recherches en agronomie sur la nutrition minérale et la fertilisation des cultures n’ont pu aborder qu’une seule face du système : l’optimisation de la production agricole par la fertilisation, mais se trouvent incapables d’analyser les deux faces du problème : la production et l’environnement.

Figure 6 : Représentation de la réponse de la culture à l’état nutritionnel azoté de la culture (INN) qui devient un invariant. L’INN obtenu par une culture est un indicateur non seulement de la capacité qu’a la culture de satisfaire son besoin en N, mais de la disponibilité globale de N à circuler entre les différentes composantes du système sol – plante – microbes, et donc des risques environnementaux afférents. Le diagnostic INN de la culture est donc aussi un diagnostic de risque environnemental qui permet de mettre en balance les deux termes du compromis production-environnement.
L’approche de diagnostic de nutrition minérale des cultures, parce que basée sur des propriétés émergentes résultant des facultés d’auto-adaptation du système sol – plante – microbiome*, rend compte d’un état global interne de ce système duquel on peut inférer aussi bien des niveaux de production possibles d’une culture que des risques environnementaux encourus, compte tenu du niveau de nutrition azotée (INN) de cette culture. Il est donc possible de prendre en compte le compromis à trouver entre les deux objectifs de production agricole et de préservation de l’environnement. En effet, un diagnostic de nutrition en azote ou phosphore d’une culture nous renseigne non seulement sur son niveau de satisfaction en ces deux éléments, et donc de son niveau de rendement possible, mais encore sur la facilité de circulation de ces éléments entre les éléments du système sol – plante – microbes - matières organiques, en d’autres mots, du degré de couplage ou de découplage de ces éléments avec le carbone du sol (Rumpel & Chabbi, 2019 ; Recous et al., 2019), et donc sur les risques potentiels d’émission dans le milieu.
Ainsi on propose de renverser totalement l’approche de la nutrition minérale et de la fertilisation des cultures. Le paradigme ancien consistait à étudier la réponse des cultures à un apport croissant d’élément fertilisant conformément au schéma de la figure 4, dans lequel la culture témoin, c’est-à-dire sans apport d’azote, s’avère soumise à la plus grande source de variabilité, celle-ci étant due à la fois au sol et au climat. La réponse à l’apport de fertilisant ne peut donc pas être interprétée de manière fonctionnelle et reste donc soumise à un grand degré d’incertitude due à l’absence de témoin « invariant ». Le paradigme nouveau au contraire prend comme témoin la culture non-limitée en azote, et grâce au diagnostic de nutrition azotée de la culture in situ (INN) il est possible d’établir une loi ayant une valeur générique quantifiant la réponse, non plus aux apports croissants de N, mais à l’intensité de déficit de nutrition N de la culture correspondant à la valeur 1-INN (Figure 6). Ce renversement de paradigme permet alors d’incorporer les variables contextuelles locales dans la valeur de l’INN observée in situ, et de faire alors ressortir une loi plus générique de réponse d’une culture au niveau de déficit en N qu’elle subit réellement dans les conditions spatiales et temporelles dans lesquelles ce diagnostic a été réalisé. De plus, ce diagnostic étant basé sur un état du peuplement végétal dans son milieu réel, il prend en compte l’ensemble des interactions entre les trois composantes du système, la plante, le sol et le microbiome, et on est en droit d’en inférer un comportement global de ce système qui doit pouvoir être traduit en termes de risques environnementaux.
Conclusion
Ainsi un changement de paradigme devient possible. D’une approche linéaire de pronostic de réponse des cultures à des apports exogènes d’éléments nutritifs lestée d’une grande incertitude pour son application contextuelle locale, il est possible de passer à une approche de diagnostic local dont la signification générique est étayée par un corpus théorique cohérent. Du coup, ce changement de paradigme permet d’effacer le niveau d’incertitude lié au pronostic, grâce à l’aptitude du diagnostic in situ à rendre compte de la diversité des contextes. La possibilité de répétition spatiale et temporelle des diagnostics in situ permet une adéquation plus étroite des décisions au contexte local. C’est ainsi que le couplage entre diagnostic in situ en temps réel et les outils de décision de fertilisation des cultures grâce à la télédétection dans le cadre de l’agriculture de précision ouvre techniquement la porte à des pratiques essentiellement correctives, à la fois dynamiques et plus précises, permettant d’assurer le maintien du statut nutritionnel des cultures à un niveau jugé à la fois optimum pour les objectifs de production et compatible avec l’environnement. Cependant, les recherches doivent se concentrer justement sur les termes de ce compromis entre optimum de production et impacts environnementaux dont les poids respectifs sont variables en fonction non seulement des contextes agronomiques et écologiques, mais aus1si économiques et sociopolitiques.
Note
[1] Les mots suivis d’un astérisque sont définis dans le glossaire en fin de texte.
Glossaire
Allométrie : en biologie du développement, l’allométrie est l’analyse des croissances relatives entre les organes, tissus ou processus d’un organisme vivant.
C3-C4 (espèces en…) : classement des espèces en fonction de leur type de métabolisme photosynthétique. Les espèces en C3 correspondent à l’ensemble des espèces cultivées tempérées y compris le riz ; les espèces en C4 correspondent essentiellement aux graminées tropicales : maïs, sorgho, canne à sucre. Leur différence de métabolisme leur confère des teneurs critiques en azote différentes, plus élevées pour les C3 que pour les C4.
Co-limitation : limitation de la croissance d’un organisme ou d’une population d’organismes par l’action simultanée d’au moins deux facteurs différents. Ce concept s’oppose à la Loi de Liebig des facteurs limitant, dans la mesure où celle-ci n’envisage pas d’actions simultanées, mais des facteurs limitants agissant les uns à la suite des autres.
Écologie fonctionnelle : discipline de l’écologie scientifique, elle étudie le rôle et les fonctions que les individus, les populations, les espèces jouent dans leur biocénose, c’est-à-dire leur communauté et leur environnement.
Générique : se dit d’une relation, d’un processus ou d’une fonction qui, observée ou étudiée dans un contexte donné, est transposable à une grande diversité de contextes.
Microbiome (du sol) : c’est l’entité formée par l’ensemble des espèces microbienne du sol (microbiote) et leur « aire de vie » dans un sol.
Paradigmes : ce sont les principes et méthodes qui fondent la recherche dans une discipline scientifique déterminée.
Propriétés émergentes (d’un système) : il s’agit des propriétés attachées à un système complexe qui ne peuvent s’exprimer qu’à l’échelle du système entier et dont la signification disparaît aux échelles d’organisation inférieures.
Réflectance (d’un couvert végétal) : ce sont les propriétés optiques des feuilles des végétaux et plus particulièrement des couverts végétaux qui modifient le spectre de la lumière réfléchie. La mesure de cette réflectance dans différentes gammes de longueurs d’onde par télédétection (voir ce terme) permet d’en déduire certaines informations sur le contenu en azote du couvert végétal.
Stoechiométrie : calcul qui permet d’analyser les quantités relatives de réactifs et de produits qui sont en jeu au cours d’une réaction chimique. Cette approche permet d’analyser les rapports entre les éléments chimiques rentrant en jeu dans la constitution des organismes vivants et le fonctionnement des écosystèmes.
Télédétection : dans ce cadre précis, il s’agit de procédés de mesure et d’interprétation des ondes électromagnétiques (principalement la lumière) émis ou réfléchis par les couverts végétaux, que ce soit au moyen d’appareillages proches portables, ou aéroportés, ou satellitaires, permettant d’en inférer des variables indicatrices de leur fonctionnement.
Références
Agren G.I. 2004. The C : N : P stoichiometry of autotrophs—theory and observations. Ecology Letters, 7 : 185-191.
Barret F., Fourty T. 1994. Radiometric estimate of nitrogen status of leaves and canopies. In (Lemaire G., ed): Diagnosis of the nitrogen status in crops. Springer, Heidelberg, pp 201-227.
Blackman F.F., 1905. Optima and limiting factors. Ann. Bot. 19, 281–295.
Bloom A.J., Chapin F.S., Mooney H.A., 1985. Resource limitations in plants – An economic analogy. Ann. Rev. Ecol. Syst., 16 : 363-92.
Blum J.J. 1977. On the geometry of four-dimensions and the relationship between metabolism and body mass. Journal of Theoretical Biology, 64 : 599-601.
Boussingault J.B., 1855. Recherches sur la végétation. De l’action du salpêtre sur le développement des plantes. Journal de Pharmacie et de Chimie, 3e série, 25 : 122-131.
Bramley R.G.V., Lawes R.A., Cook S.E., 2013. Spatially distributed experimentation. In : Oliver M.A., Bishop T., Marchant B. (eds.), Precision Agriculture for Sustainability and Environmental Protection. Routledge, pp. 205-218. doi.org/10.4324/9780203128329.
Briat J.F., Rouached H., Tissot N., Gaymard F., Dubos C. 2015. Integration of P, S, Fe, and Zn nutrition signals in Arabidopsis thaliana : potential involvement of Phosphate Starvation Response 1 (PHR1). Front Plant Sci., 28 (6) : 290.
Briat J.F. Gojon A., Rouached H., Plassard C., Lemaire G., 2020. Reappraisal of the concept of nutrient availability for plants in soils at the light of the recent molecular physiology advances. European Journal of Agronomy (submitted).
Caloin M., Yu O., 1984. Analysis of the time course change in nitrogen content of Dactylis glomerata L. using a model of plant growth. Ann. Bot., 54 : 69-76.
Cao Q., Miao Y.X., Shen J.N., Yu W.F., Yuan F., Cheng S.S., Huang S.Y., Wang H.Y., Yang W., Liu F.Y., 2016. Improving in-season estimation of rice yield potential and responsiveness to topdressing nitrogen application with Crop Circle active crop canopy sensor. Precis. Agric., 17: 136-154. doi.org/10.1007/s11119-015-9412-y.
Cornforth I.S.; Steele K.W. 1981. Interpretation of maize leaf analysis in New Zealand. NZ journal of experimental agriculture, 9 : 91-96.
Cossani C.M., Sadras V.O., 2018. Water-nitrogen co-limitation in grain crops. Adv. Agron., 150 : 231–274.
Devienne-Barret F., Justes E., Machet J.M., Mary B., 2000. Integrated control of nitrate uptake by crop growth rate and soil nitrate availability under field conditions. Ann. Bot., 86 : 995-1005. doi : 10.1006/anbo.2000.1264
De Wit C.T., 1994. Resource use analysis in agriculture: a struggle for interdisciplinarity. In : Fresco L.O., Stroosnijder L., Bouma J., van Keulen H. (eds.), The Future of the Land : Mobilizing and Integrating Knowledge of Land Use Options. Wiley, NewYork, pp. 41-55.
Duru M., Thellier L., 1997. N and P-K status of herbage: use for diagnosis of grasslands. In: Lemaire G., Barns I., eds, Diagnostic procedures for crop N management and decision making. Science Update, INRA Editions, Paris.
Galloway J.N., Cowling E.B., 2002. Reactive nitrogen and the world: 200 years of change. Ambio 3 : 64-71.
Greenwood D.J, Lemaire G., Gosse G., Cruz P., Draycott A., Neeteson J.J., 1990. Decline in percentage N of C3 and C4 crops with increasing plant mass. Ann. Bot., 66 : 425-436.
Hanks J., Ritchie J.T. (eds.), 1991. Modelling Plant and Soil Systems. Agronomy Monograph No. 31, American Society of Agronomy, Madison, 545 p.
Hébert J., 1969. La fumure azotée du blé tendre d’hiver. Bulletin Technique d'Information 244 : 755-766.
Kho R.M., 2000. On crop production and the balance of available resources. Agriculture, Ecosystems and Environment 80: 71-85
Lejay L., Tillard P., Lepetit M., 1999. Molecular and functional regulation of two nitrate uptake systems by N and C-status of Arabidopsis plants. Plant J., 18: 509-519.
Lemaire G., Salette, J. 1984. Relation entre dynamique de croissance et dynamique de prélèvement d’azote pour un peuplement de graminées fourragères. I - Etude de l’effet du milieu. Agronomie, 4 (5) : 423-430.
Lemaire G., Cruz P., Gosse G., Chartier M., 1985. Étude des relations entre la dynamique de prélèvement d’azote et la dynamique croissance en matière sèche d’un peuplement de luzerne (Medicago sativa L.). Agronomie, 5 : 685-692.
Lemaire G., Jeuffroy M.H., Gastal F., 2008. Diagnosis tool for plant and crop N status in vegetative stage. Theory and practices for crop N management. Eur. J. Agron., 28 : 614-624. doi : 10.1016/j.eja.2008.01.005
Lemaire G., van Oosterom E., Sheehy J., Jeuffroy M.H., Massignam A., Rossato L., 2007. Is crop demand more closely related to dry matter accumulation of leaf area expansion during vegetative growth? Field Crops Res., 100 : 91-106. https : //doi.org/10.1016/j.fcr.2006.05.009
Lemaire G., 2019. Allometric approach to crop nutrition and implications for crop diagnosis and phenotyping. A review. Agronomy for Sustainable Development (in press).
Liebig J. von, 1855. Die Grundsätze der Agricultur-Chemie mit Rücksicht auf die in England angestellen Untersuchungen. 2 Auflage, Braunschweig, 107 p.
Liebscher G., 1895. Untersuchungen über die Bestimmung des Düngerbedürfnisses der Ackerböden und Kulturpflanzen. J. Landwirtsch., 43: 49-216.
Machet J-M., Dubrulle P., Damay N., Duval R., Julien J.-L., Recous S., 2017. A Dynamic Decision-Making Tool for Calculating the Optimal Rates of N Application for 40 Annual Crops while Minimising the Residual Level of Mineral N at Harvest. Agronomy, 7, 73; doi : 10.3390/agronomy7040073
Meynard J.M., Justes E., Machet J.M., Recous S., 1997. Fertilisation azotée des cultures annuelles de plein champ. In : Lemaire G., Nicolardot B. (eds), Maîtrise de l’azote dans les Agrosystèmes. INRA Paris, France, Les Colloques de l'INRA, Vol. 83 : 183-200.
Meynard J-M., Cerf M., Guichard L., Jeuffroy M-H., Makowski D., 2002. Which decision support tools for the environmental management of nitrogen? Agronomie 22 : 817-829. DOI : 10.1051/agro : 2002064
Mitscherlich E.A., 1924. Die Bestimung des Düngerbedürfnisses des Bodens. Paul Parey, Berlin, 100 p.
Monteith J.L., 1994. Principles of resource capture by crop stands. In : Monteith J.L., Scott R.K., Unsworth M.H. (eds.), Resource Capture by Crops. Proc. of Easter Schools in Agricultural Science, Nottingham University Press, Nottingham, pp. 1-15.
Niklas K.J., Owens T., Reich P.B., Cobb E.D., 2005. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth. Ecol. Lett., 8: 636–642.
O’Brien J.A., Vega A., Bouguyon E., Krouk G., Gojon A., Coruzzi G., Gutierrez R.A. 2016 Nitrate transport, sensing and responses in plants. Molecular Plant, 9 : 837-856.
Ohkubo Y., Tanaka M., Tabata R., Ogawa-Ohnishi M., Matsubayashi Y., 2017 Shoot-to-root mobile polypeptides involved in systemic regulation of nitrogen acquisition. Nature Plants, 3 : 17029
Rabinovitch E.L., 1951. Photosynthesis and Related Processes. Vol. II, Part 1. Interscience Publishers, New York, pp. 858-885.
Ratjen A.M., Lemaire G., Kage H., Plénet D., Justes E., 2018. Key variables for simulating leaf area and N status : biomass based relations versus phenology driven approaches. Eur. J. Agron., 100 : 110-117. doi : 10.1016/j.eja.2018.04.008
Ravier C., Jeuffroy M.H., Meynard J.M., 2016. Mismatch between a science-based decision tool and its use : The case of the balance-sheet method for nitrogen fertilization in France. NAJS-Wageningen Journal of Life Science 79 : 31-40. doi.org/10.1016/j.njas.2016.10.001.
Ravier C., Jeuffroy, M-H., Gate P., Cohan J-P., 2018. Combining user involvement with innovative design to develop a radical new method for managing N fertilization. Nutr. Cycl. Agroecosyst. 20 (110) : 117-134. doi.org/10.1007/s10705-017-9891-5.
Recous, S., Lashermes G., Bertrand I., Duru M., Pellerin M., 2019. C-N-P decoupling processes linked to arable cropping management systems in relation with intensification of production. In : Lemaire G, de Facio Carvalho P.C., Kronberg S., Recous S. (eds) Agrosystem diversity. Reconciling contemporary agriculture and environment. Elsevier, Academic Press.
Reich P.B., Oleksyn J., Wright I.J., Niklas K.J., Hedin L., Elser J.J., 2018. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes. Proc. R. Soc. B., 277 : 877-883. doi : 10.1098/rspb.2009.1818.
Rémy J.C.; Hébert J. 1977. Le devenir des engrais azotés dans le sol. C. R. Acad. Agric. Fr., 63: 700-714
Ringeval B., Augusto L., Monod H., van Apeldorn D., Bouwman L., Yang X., Achat D.L., Chini L.P., van Oost K., Guenet B., Wang R., Decharme B., Nesme T., Pellerin S., 2017. Phosphorus in agricultural soils : drivers of its distribution at the global scale. Global Change Biology, doi : 10.1111/gcb.13618.
Rumpel C., Chabbi A., 2019. Plant-Soil interactions control CNP coupling and decoupling processes in agroecosystems with perenniem vegetation. In : Lemaire G, de Facio Carvalho P.C., Kronberg S., Recous S. (eds), Agrosystem diversity. Reconciling contemporary agriculture and environment. Elsevier, Academic Press. ISBN : 978-0-12-811050-8.
Salette J, Huché L (1991) Diagnostic de 1'état de nutrition minérale d'une prairie par l'analyse minérale du végétal: principes, mise en oeuvre, exemples. Fourrages 125, 3–18.
Sinclair T.R., Park W.I., 1993. Inadequacy of the Liebig limiting-factor paradigm for explaining varying crop yields. Agron. J., 85 : 742–746
Smith P.F., 1962. Mineral analysis of plant tissues. Ann. Rev. Plant Physio., 13 : 81-108.
Soussana J.F., Lemaire G., 2014. Coupling C and N cycles for environmentally sustainable intensificationof grasslands and crop-livestock systems. Agriculture, Ecosystem & Environment, 190: 9-17.
Tilman D., Cassman K.G., Matson P.A., Naylor R., Polasky S., 2002. Agricultural sustainability and intensive production practices. Nature, 418 (6898) : 671-677.
Ulrich A., 1952. Physiological bases for assessing the nutritional requirements of plants. Ann. Rev. Plant Physiol,. 3, 207-228.
Valkama ER, Uusitalo R, Turtola E, 2011. Yield response models to phosphorus application : A research of Finnish field trials to optimize P use of cereals. Nutr. Cycling Agroecosyst., 91 : 1-15.
Walworth J.L., Sumner, M.E., 1987. The Diagnosis and Recommendation Integrated System (DRIS). Advances in soil science, 6 : 149-188.
Ziadi N., Bélanger G., Cambouris A., Tremblay N., Nolin M.C., Claessens A., 2007. Relationship between P and N concentration in corn. Agron. J., 99 : 833-841. doi : 10.2134/agronj2006.0199.
Ziadi N., Bélanger G., Cambouris A., Tremblay N., Nolin M.C., Claessens A., 2008. Relationship between phosphorus and nitrogen concentrations in spring wheat. Agron. J., 100 : 80-86. doi : 10.2134/agronj2007.0119.

Les articles sont publiés sous la licence Creative Commons (CC BY-NC-ND 2.0)
Pour la citation et la reproduction de cet article, mentionner obligatoirement le titre de l'article, le nom de tous les auteurs, la mention de sa publication dans la revue AES et de son URL, la date de publication.