

Effets à court terme des digestats bruts de méthanisation sur la productivité et la qualité fourragère ainsi que sur le fonctionnement microbien sous prairies
LAFLOTTE Alexandre1, PACAUD Stéphane1, DAIGNEUX Benjamin2, POIRRIER Fanny3, GENESTIER Julie3, RAVARD Benjamin1, LE ROUX Yves4, PIUTTI Séverine3
1Université de Lorraine, ENSAIA, Centre R&D, TSA 40602, F-54518 Vandoeuvre-lès-Nancy Cedex alexandre.laflotte@univ-lorraine.fr
2Agra-Ost, Klosterstrasse 38, B-4780 Sankt-Vith
3Université de Lorraine, INRAE, UMR 1121 Agronomie et Environnement, TSA 40602, F-54518 Vandoeuvre-lès-Nancy Cedex
4Université de Lorraine, UR-AFPA, TSA 40602, F-54518 Vandoeuvre-lès-Nancy Cedex
Résumé
Dans le cadre d’un projet INTERREG, cinq sites expérimentaux de prairies permanentes sont suivis depuis 2017 avec pour objectif de comparer dans différentes conditions pédoclimatiques les effets de différentes modalités de fertilisation (digestat de méthanisation vs engrais de synthèse vs lisier brut vs témoin non fertilisé).
Cet article montre que 2 ans après le début des applications des différents fertilisants -à unités d’azote équivalentes- les rendements en biomasse sont significativement plus élevés sur des parcelles fertilisées par du nitrate d’ammonium en comparaison à des parcelles non fertilisées. Les rendements en biomasse des parcelles fertilisées par du lisier ou du digestat sont intermédiaires. Cependant les valeurs fourragères et les teneurs en éléments minéraux ne permettent pas de discriminer les modalités de fertilisation. De même, à ce stade, il n’est pas possible de distinguer d’effet significatif des modalités de fertilisation sur le fonctionnement microbien des sols.
Mots-clés : Digestat de méthanisation, rendement, qualité fourragère, variables microbiennes, Interreg Grande Region
Abstract
Early effects of digestates on the productivity and quality of forages and on soil microbial variables in grasslands
In a European Project gathering french, belgian, luxembourgish and german partners, five permanent grassland experimental sites have been monitored since 2017. The aim of this project is to compare the effects of different fertilizers under different pedoclimatic conditions (digestate vs chemical fertilizer vs manure vs unfertilized control).
This article shows that 2 years after the start of the application of the different fertilizers – for the same nitrogen units - the biomass yields are significantly higher on plots fertilized with ammonium nitrate compared to unfertilized plots. The biomass yields of plots fertilized with manure or digestate are intermediate. However, the feeding values and the mineral contents do not make it possible to identify the fertilizer. Likewise, after two years, it is not possible to distinguish a significant effect of the modes of fertilizer on the soil microbial variables.
Keywords : digestate, yield, feeding value, soil microbial variables, Interreg Grande Region
Introduction
L’utilisation de biomasse en méthanisation permet de générer des coproduits de réaction correspondant à une matrice organique résiduelle i.e. digestats bruts (Zhang et al., 2007) qui peuvent être utilisés en agriculture comme engrais et/ou amendements organiques (Möller et Müller, 2012). De nombreux verrous ont été levés ces dernières années sur la faisabilité technique autour de la mise en place d’unités de méthanisation, mais de nombreuses questions sont actuellement adressées à la profession agricole concernant l’épandage au champ et la valorisation agronomique des digestats bruts. Les digestats pourraient ainsi améliorer (i) à très court terme la production agricole (biofertilisant), (ii) à plus long terme la fertilité des sols agricoles (activateurs de la vie du sol), les deux potentialités devant permettre de diminuer la dépendance des systèmes agricoles vis-à-vis des intrants de synthèse.
Concernant les aspects de biofertilisation, la caractéristique essentielle des digestats à considérer est sa forte disponibilité en éléments minéraux, notamment en azote avec en une forme ammoniacale pouvant représenter entre 44 et 81% de l’azote total (Möller et Müller, 2012). Ces digestats peuvent ainsi dans certaines conditions et pour certaines espèces végétales remplacer les engrais de synthèse (Alburquerque et al., 2012 ; Coelho et al., 2019). Dans le cas des espèces prairiales et notamment du ray-grass anglais, si les digestats peuvent significativement augmenter le rendement en comparaison d’un témoin non fertilisé, leur valeur fertilisante reste néanmoins souvent inférieure aux engrais azotés de synthèse (Walsh et al., 2012). Néanmoins, la majorité des résultats disponibles dans la littérature scientifique proviennent d’essais en conditions contrôlées réalisées en mésocosmes sur des cycles de développement courts.
Les matières organiques des sols (MOS) sont le pilier de la fertilité des sols (Manlay et al., 2007), définie comme la capacité à « fournir des nutriments essentiels pour la croissance des plantes cultivées, soutenir une communauté biotique diversifiée et active et présenter une structure favorable » (Mäder et al., 2002). Ces MOS constituent notamment à la fois un réservoir de nutriments pour les plantes dans les agroécosystèmes et une source de carbone et d’énergie pour les micro-organismes du sol, acteurs majeurs de la minéralisation des MOS.
Les conditions pédoclimatiques et les pratiques culturales influencent la taille et l’activité des micro-organismes du sol. Les pratiques agricoles permettant d’augmenter les entrées de carbone dans le sol sont de manière générale favorables au fonctionnement microbien. Ainsi, l’épandage régulier d’effluents d’élevage permet un retour au sol de substrats organiques pouvant stimuler les communautés microbiennes tant en termes de croissance que d’activité de minéralisation des MOS (Zhang et al., 2012 ; Zhang et al., 2015). Aussi les digestats, du fait de caractéristiques différentes (ratio C/N faible, forte teneur en azote minéral, C moins labile…) par rapport à des effluents bruts (lisier ou fumier) pourraient modifier à court et moyen terme le fonctionnement microbien des sols.
Initié en 2016, le projet Interreg PERSEPHONE a notamment pour objectif de comparer dans différentes conditions pédoclimatiques (France, Belgique, Luxembourg et Allemagne) les effets de différentes modalités de fertilisation (digestat vs engrais chimiques vs lisier brut vs témoin non fertilisé) sur la production de fourrage en prairies (rendement et qualité fourragère) et sur des variables microbiennes (biomasse microbienne carbonée et azotée ainsi que des mesures d’activités enzymatiques relatives aux différents cycles biogéochimiques du carbone, azote, phosphore et soufre) en lien avec la fertilité biologique des sols (Petitjean et al., 2019).
Matériel et méthodes
Les sites expérimentaux

Figure 1 : Carte des différents sites expérimentaux suivis entre 2017 et 2019
Cinq sites expérimentaux sont suivis dans le cadre de ce projet Interreg (Figure 1). Les sites d’Emmels, Steinborn, Erpeldange et La Bouzule ont été mis en place au début du projet en février 2017. Le site de Grendel qui était déjà installé et pour lequel nous n’avons pas de caractérisation initiale n’a pas été inclus dans l’analyse des résultats de fonctionnement microbien. Les caractéristiques physico-chimiques des sols des 4 sites retenus sont présentés dans le Tableau 1. Le tableau 1 met en évidence des textures différentes selon les sites expérimentaux considérés, avec le site de Steinborn présentant un sol à dominante sableuse alors que les sites d’Emmels et Erpeldange sont à dominante limoneuse. Le site de la Bouzule présente une teneur en calcaire plus importante et un pH proche de 8,2 contre 5,8 sur le site d’Emmels. La teneur en C organique est deux fois plus importante sur le site de Emmels (38,4‰) que de Steinborn (19,5‰). Les sites de la Bouzule et d’Erpeldange ont des teneurs en C organique intermédiaire proches autour de 32‰. Cette caractérisation initiale met en évidence des différences de propriétés physico-chimiques plus importantes entre les sites qu’entre les blocs d’un même site. Cela suggère que nous disposons donc d’un gradient pédoclimatique et que les différents sites présentent une faible hétérogénéité.
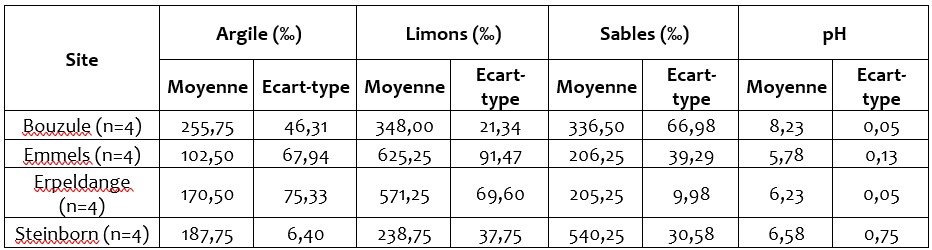
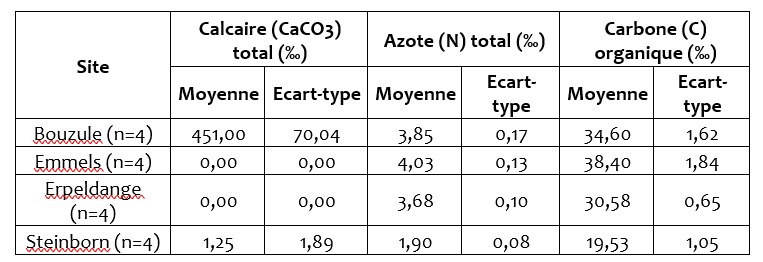
Tableau 1 : Principales caractéristiques physico-chimiques des sols présents sur les sites au moment de la mise en place des modalités de fertilisation.
Modalités étudiées
L’ensemble des sites correspondent à des prairies permanentes (PP) à l’exception du site de La Bouzule qui présente à la fois un essai sur PP et un autre sur prairie temporaire (PT) implantée en 2017. Cinq modalités de fertilisation communes à tous les sites ont été mises en place selon un dispositif en bloc avec randomisation (4 répétitions / modalité) pour l’étude :
- Témoin non fertilisé (Contrôle)
- Digestat brut de la ferme du Faascht (DBF) (commun à tous les sites) : 230 unités N
- Digestat brut local (DBL) : 230 unités N
- Lisier brut local (LBL) : 230 unités N
- Nitrate d’ammonium (Nammo) : 230 unités N
Les apports de fertilisants ont été fractionnées en 4 apports. Les 230 unités N ont été réparties équitablement sur ces 4 apports. Le tableau 2 précise les dates d’apport pour chacune des années.

Tableau 2 : Date d’apport des fertilisants en 2017 et 2018
Chaque modalité correspond à une microparcelle de 9 à 12m2 selon les sites. Les caractéristiques des différents digestats sont détaillées dans le tableau 3.

Tableau 3 : Caractéristiques des différents digestats et lisiers selon les sites d’épandage.
Variables mesurées
Production fourragère (quantité de biomasse et qualité fourragère)
Les campagnes de fauche 2017 et 2018 ont été analysées sur l’ensemble des sites (Grendel-Attert, Bouzule PP, Erpeldange, Steinborn, Emmels). Trois fauches ont été réalisées par an et par site (sauf Bouzule Prairie Permanente (PP) 2018 – 2 fauches). Pour la production de biomasse, une pesée de la biomasse totale fauchée sur chaque microparcelle est réalisée à chaque fauche. La détermination de la teneur en MS est faite sur un échantillon par microparcelle placé en étuve à 65 °C.
Pour ce qui est de la qualité fourragère, le second essai présent sur le site de La Bouzule (PT) a été intégré à l’analyse. Les échantillons ainsi séchés sont envoyés pour analyse de teneur en minéraux et valeur fourragère :
- Les minéraux analysés sont MgO, CaO, Na2O, K2O, P2O5 (respectivement Magnésium, Calcium, Sodium, Potassium et Phosphore), cendres
- Les indicateurs de valeurs fourragères sont MAT, UFL, FOS, ADL, ADF, NDF, CELL, PROTTOT, PDIE, PBD, SSTOTAUX (respectivement Matière Azotée Totale, Unité fourragère Lait, Matière organique fermentescible, Acid Detergent Lignin, Acid Detergent Fiber, Neutral Detergent Fiber, Cellulose, Protéines totales, Protéines digestibles dans l’intestin grêle permises par l’énergie, Protéines brutes digestibles, Sucres Solubles totaux)
Variables microbiennes
Des prélèvements sont effectués deux fois par an : en février / mars avant les premiers apports d’azote et en octobre avant la dernière fauche de l’année. Quatre prélèvements sont réalisés sur chaque micro-parcelle sur 15 cm de profondeur. Les sols sont ensuite tamisés à 5 mm et stockés à 4°C quelques jours avant détermination des différentes variables microbiennes. Pour caractériser le fonctionnement du sol, nous avons suivi différents indicateurs :
- La biomasse microbienne carbonée et azotée. La méthode utilisée pour le dosage de la biomasse microbienne du sol est celle de la fumigation-extraction (Vance et al., 1987).
- Six activités enzymatiques microbiennes impliquées dans différentes étapes de la minéralisation des MOS. : Les leucine aminopeptidases (hydrolyse des liaisons peptidiques des peptides), Nacetylglucosaminidases (hydrolyse de la chitine), les ß-glucosidases (hydrolyse des oligosaccharides), les arylsulfatases (hydrolyse des esters de sulfates) et les phosphatases acides (hydrolyse des esters de phosphate). Ces activités enzymatiques (exprimées en nmol/h/g sol sec) ont été dosées par fluorimétrie (Petitjean et al., 2018).
- les pools extractibles à l’eau chaude de carbone et azote total (Sparling et al., 1998).
En raison de l’antériorité du site d’Attert-Grendel, ces variables n’ont été mesurées que sur les sites de Emmels, Steinborn, Bouzule PP et PT et Erpeldange en mars 2019, permettant ainsi de mesurer les effets de deux années de pratiques culturales.
Analyse statistique
Pour la biomasse produite, une analyse de variance à deux facteurs (Site et Traitement) a été réalisée pour chaque année. Pour la qualité des fourrages (teneur en minéraux et valeurs fourragères), une analyse de données non-supervisée a été réalisée avec une analyse en composante principale (ACP) suivie d’une classification ascendante hiérarchique (CAH). Les teneurs en MgO, CaO, Na2O, K2O, P2O5, cendres ont été utilisées en variables actives et les rendements en MS et les dates de fauche ont été notés en variables illustratives.
Pour les valeurs fourragères, MAT, UFL, FOS, ADL, ADF, NDF, CELL, PROTTOT, PDIE, PBD, SSTOTAUX ont été utilisées en variables actives alors que les rendements en MS et les dates de fauche ont été utilisés en variables illustratives.
Résultats et discussion
Quantité de biomasse produite en 2017 et 2018
Les quantités de biomasse (kg MS ha-1) produites par coupe ont été cumulées afin de comparer les 5 modalités de fertilisation entre elles (Figures 2 à 5). Pour les deux années malgré une interaction traitement*site (p=0,06 en 2017 et p=0,008 en 2018) des tendances peuvent être observées : la modalité Nammo est celle qui permet d’obtenir la biomasse la plus élevée sur les deux années, et à l’inverse la modalité Contrôle correspond au rendement le plus faible. Pour les digestats et le lisier, aucune hiérarchie entre ces trois modalités ne peut être établie (Figure 2 et Figure 3). L’analyse de données met en évidence un fort effet site avec des rendements moyens notamment sur le site d’Erpeldange quasiment trois fois plus élevés que sur le site d’Attert-Grendel.
Ainsi, malgré cette interaction traitement*site, une analyse de variance testant uniquement l’effet traitement a été réalisée (Figure 4 et Figure 5) ; comme déjà vu sur les figures 2 et 3 quelle que soit l’année, la modalité Nammo permet d’obtenir le niveau de biomasse produite le plus élevée en comparaison de la modalité Contrôle qui présente le niveau de biomasse le plus faible. Les digestats et le lisier sont intermédiaires.




La biomasse obtenue en 2017 par la fertilisation au nitrate d’ammonium est significativement plus élevée (7190 kg MS ha-1) que celle permise par le témoin non fertilisé (4880 MS ha-1), soit 2310 kg MS ha-1 en plus (+47%) avec 230 unités d’azote sous forme de nitrate d’ammonium – en moyenne sur tous les sites – soit un peu plus de 10 kg MS ha-1 unité N-1 (Figure 4). Les trois autres modalités Digestat Brut Local, Digestat Brut du Faascht, Lisier Brut Local sont intermédiaires. Elles ne sont pas différentes entre elles mais sont statistiquement différentes des modalités NAmmo et Contrôle. Ainsi à l’issue de la première année de façon globalisée sur l’ensemble des sites, les modalités digestats et lisier ne se discriminent pas sur la base du rendement fourrager.
En 2018, le rendement de la modalité NAmmo est statistiquement supérieur aux modalités Digestat brut local (DBL) et Contrôle (Figure 5). Les modalités Digestat Brut de Faascht (DBF) et Lisier (LBL) sont intermédiaires et non différentes des modalités NAmmo et DBL. Si l’on compare les rendements 2017 et 2018, il apparait que :
- Les rendements moyens 2018 de chaque modalité sont plus élevés que l’année précédente (+15 à +25% selon les modalités)
- Le classement des modalités par ordre de productivité décroissante est le même en 2018 qu’en 2017, avec toujours une productivité plus élevée pour NAmmo, puis les deux digestats (DBL et DBF) et le lisier (LBL) et enfin le Contrôle.
- L’apport de 230 unités N sous forme de nitrate d’ammonium permet en 2018 la production de 2170 kg MS ha-1 supplémentaire par rapport au contrôle, proche du résultat observé en 2017.
- L’effet site est identique en 2017 et 2018.
Qualité des fourrages récoltés : teneur en minéraux
L’évolution des variations de la teneur en minéraux des fourrages a été abordée par une analyse de variance à trois facteurs. Cette approche permet de mettre en évidence systématiquement des effets simples significatifs (site, traitement, coupe), des interactions doubles variables en fonction des minéraux testés mais pas d’interaction triple significative. Pour des questions de lisibilité le tableau d’analyse de variance à trois facteurs Site*Traitement*Coupe n’est pas présenté.
Les histogrammes présentés en figure 6A, 6B, 6C, 6D illustrent pour deux teneurs en minéraux (K2O et P2O5) et uniquement pour la première coupe de chaque année, les interactions Traitement*Site. Il existe également une interaction Traitement*Site différente selon les années pour un élément considéré.
Ainsi il apparaît difficile par l’ANOVA de donner des conclusions génériques sur l’effet des facteurs simples ou des interactions doubles ou triple pour les minéraux étudiés.

Figure 6 : Teneur des fourrages récoltés en mg pour 100g sec de K2O (A et B) et P2O5 (C et D) pour chaque traitement et chaque site de la première coupe de 2017 (A et C) et 2018 (B et D).
Une analyse en composantes principales (ACP) est réalisée sur les 6 variables qui caractérisent les teneurs en minéraux des fourrages. Les deux premières dimensions de l’ACP expliquent environ 60% de l’inertie du nuage de points. Les variables MGO, CAO, NA2O et K2O (respectivement teneur en magnésium, calcium, sodium et potassium) contribuent le plus à la construction de l’axe 1, et les variables cendre, P2O5 et K2O contribuent le plus à la construction de l’axe 2. Une projection (Figure 7A) des individus selon leur appartenance aux « modalité de fertilisation », variable qui ne participe pas à la construction de l’ACP, montre qu’il n’est pas possible de mettre en évidence par cette méthode d’effet des modalités de fertilisation sur la teneur en minéraux des fourrages récoltés. Les mêmes conclusions peuvent être proposées pour le lien entre le rendement en matière sèche et la date utilisées en variables illustratives quantitatives et les 6 variables qui caractérisent les teneurs en minéraux des fourrages (Figure 7B).

Figure 7 : Analyse en composantes principales (dimensions 1 et 2) effectuée sur les teneurs en minéraux mesurées sur les 5 modalités de fertilisation pour l’ensemble des sites en 2017 et 2018. Les individus sont regroupés sur le graphique A selon la variable « modalité de fertilisation » et les variables de construction du plan factoriel sont représentées sur le cercle des corrélations sur le graphique B.
La classification ascendante hiérarchique (CAH) réalisée (données non montrées) confirme comme la représentation graphique (Figure 7A) que les modalités de fertilisation n’ont pas d’effets sur les teneurs en minéraux des fourrages. En revanche, si les individus sont regroupés selon la variable « sites expérimentaux » (Figure 8A), les regroupements suggèrent un effet des conditions pédoclimatiques supérieur à l’effet du mode de fertilisation. De la même façon, le numéro d’ordre de la coupe dans l’année apparaît supérieur à l’effet du mode de fertilisation (Figure 8B).

Figure 8 : Analyse en composantes principales (dimensions 1 et 2) effectuée sur les teneurs en minéraux mesurées sur les 5 modalités de fertilisation pour l’ensemble des sites en 2017 et 2018. Les individus sont regroupés sur le graphique A selon la varia
Qualité des fourrages récoltés : valeurs fourragères
Toutes les variables relatives aux « valeurs fourragères » sur les deux années ont été utilisées pour réaliser une ACP. Les deux premières dimensions de l’ACP expliquent plus de 81 % de l’inertie du nuage de points (Figure 9B). Les variables UFL, FOS, ADL, ADF, NDF et CELL (Unité fourragère Lait, Matière organique fermentescible, Acid Detergent Lignin, Acid Detergent Fiber, Neutral Detergent Fiber, Cellulose) contribuent le plus à la construction de l’axe 1, alors que PROTTOT, PDIE, PBD, SSTOTAUX (Protéines totales, Protéines digestibles dans l’intestin grêle permises par l’énergie, Protéines brutes digestibles, Sucres Solubles totaux) contribuent le plus à la construction de l’axe 2 (Figure 9B).
Une projection (Figure 9A) des individus selon leur appartenance aux « modalité de fertilisation », variable qui ne participe pas à la construction de l’ACP, montre qu’il n’est pas possible de mettre en évidence par cette méthode d’effet des modalités de fertilisation sur la valeur fourragère des fourrages récoltés.

Figure 9 : Analyse en composantes principales (dimensions 1 et 2) effectuée sur les valeurs fourragères mesurées sur les 5 modalités de fertilisation pour l’ensemble des sites en 2017 et 2018. Les individus sont regroupés sur le graphique A selon la variable « modalité de fertilisation » et les variables de construction du plan factoriel sont représentées sur le cercle des corrélations sur le graphique B.
Comme pour la teneur des fourrages en minéraux, l’effet site et l’effet du numéro d’ordre de la fauche apparaissent plus importants que l’effet modalité de fertilisation. Les deux analyses en composantes principales (Figure 10A et 10B) montrent que les variables « site » et « Numéro d’ordre de la fauche », qui ne participent pas à la construction de l’ACP, permettent de discriminer des groupes d’individus appartenant à la même modalité (site pour la Figure 10A et numéro de fauche pour la Figure 10B).

Figure 10 : Analyse en composantes principales (dimensions 1 et 2) effectuée sur les valeurs fourragères mesurées sur les 5 modalités de fertilisation pour l’ensemble des sites en 2017 et 2018. Les individus sont regroupés sur le graphique A selon la variable « site » et sur le graphique B selon la variable illustrative « numéro d’ordre de la coupe »
En comparaison à un effluent d’élevage brut (fumier ou lisier), le digestat contient potentiellement plus d'azote biodisponible pour les plantes en raison d'une plus grande proportion des formes d'ammonium (NH4+) (Möller et Müller, 2012). Nos résultats mettent en évidence qu’en 2017, si les digestats ont un effet fertilisant significatif en comparaison d’un témoin non fertilisé, ils ne permettent pas d’atteindre une biomasse fourragère équivalente à la modalité ammonitrate et ne se différencient pas des effluents bruts. En 2018, les digestats apparaissent intermédiaires entre le témoin non fertilisé et la modalité ammonitrate. Peu d’études sont référencées dans la bibliographie sur les effets de l’épandage de digestats sur les rendements et la qualité fourragère des prairies permanentes. La majorité des essais sont conduits sur des modèles prairies temporaires avec des peuplement monospécifique de ray-grass anglais ou en mélange avec des légumineuses semés en conditions contrôlées ou sur des parcelles d’essai. De plus les digestats de méthanisation présentent une grande diversité de composition en fonction de la ration et de la conduite du procédé de méthanisation (Guilayn et al., 2019). En conditions contrôlées, Walsh et al. (2012) mettent en évidence une biomasse cumulée sur 16 semaines du mélange ray-grass-trèfle blanc significativement supérieure avec une fertilisation à base de digestats de méthanisation (150 unités d’N) en comparaison des autres modalités de fertilisation (témoin non fertilisé, effluents bruts, engrais minéral). Un essai reprenant ces mêmes modalités de fertilisation sur des prairies temporaires nouvellement semées en ray-grass entre 2011 et 2013 montre d’une part des rendements obtenus la 1ère année du même ordre de grandeur que nos essais (environ 9 tonnes de MS ha-1) et d’autre part que les rendements cumulés sur 3 ans sont significativement plus élevés pour la modalité «digestat brut» que pour la modalité «nitrate d’ammonium», ces rendements n’étant pas différents de ceux de la modalité «lisier brut» (Walsh et al., 2018). Concernant la teneur en minéraux et la qualité fourragère, aucun effet du mode de fertilisation n’a pu être mis en évidence en 2017 et 2018 ce qui est en cohérence avec les travaux de Walsh et al. (2018). Le fait que la biomasse récoltée dans notre étude présente des caractéristiques différentes selon les sites étudiés pourrait s’expliquer par des différences à la fois de paramètres physico-chimiques des sols et de pratiques de gestion (fertilisation, régime de fauche…) antérieures à la mise en place des essais qui sont certainement à l’origine de différences de compositions floristiques entre les sites. En effet des apports répétés d’azote sous forme minérale peuvent diminuer la proportion de légumineuses dans les mélanges prairiaux et la fixation symbiotique (Nesheim et al., 1990), ce qui peut modifier la qualité fourragère.
Fonctionnement microbien des sols
Le tableau 4 présente les moyennes et les écarts-types des variables microbiennes suivies sur les différents sites.

Tableau 4 : Moyenne ±écart-type des variables microbiennes des sols (0-15 cm) pour les différents dispositifs expérimentaux.
L’analyse en composantes principales (ACP) permet ici de regrouper toutes les variables mesurées sur les échantillons de sol des différents sites échantillonnés en mars 2019 (Tableau 4) afin de synthétiser les données. Cette représentation, avec les deux premières dimensions de l’ACP, explique environ 62 % de l’inertie du nuage de points (Figure 11). Le graphique B, permet de conclure que les variables HWC, HWN (pools extractibles à l’eau chaude de carbone et d’azote) BMN et BMC (carbone et azote d’origine microbienne) contribuent le plus à la construction de l’axe 1. Pour l’axe 2, ce sont les activités enzymatiques ARS et NAG respectivement en lien avec le cycle du soufre et de l’azote qui contribuent le plus à sa construction. Le graphique A qui met en évidence le positionnement des individus dans le plan factoriel déterminé par l’ensemble des variables mesurées, permet de voir que l’ensemble des individus ne sont pas situés selon les mêmes coordonnées sur les deux axes du plan factoriel. Les cercles correspondent à l’intervalle de confiance des valeurs prises par les individus appartenant à chaque modalité de fertilisation. Ces cercles se chevauchent les uns les autres ce qui suggère qu’au bout de deux années de mise en place des essais, il n’est pas encore possible de distinguer d’effet des modalités de fertilisation sur le fonctionnement microbien du sol. Néanmoins si l’on considère les sites individuellement des différenciations se dessinent notamment sur le site de Steinborn entre les modalités fertilisées et les modalités non fertilisées.

Figure 11 : Analyse en composantes principales (dimensions 1 et 2) effectuée sur les variables microbiennes mesurées sur les 5 modalités de fertilisation pour l’ensemble des sites en mars 2019. Les individus sont regroupés sur le graphique A selon la variable illustrative « modalité de fertilisation » et les variables de construction du plan factoriel sont représentées sur le cercle des corrélations sur le graphique B.
Néanmoins si l’on regroupe les individus non plus selon les modalités de fertilisation mais selon le site expérimental auxquels ils appartiennent, la Figure 12 met en évidence un regroupement significatif ce qui suggère un effet des conditions pédoclimatiques supérieures au régime de fertilisation.

Figure 12 : Analyse en composantes principales (dimensions 1 et 2) effectuée sur les variables microbiennes mesurées sur les 5 modalités de fertilisation pour l’ensemble des sites en mars 2019. Les ellipses correspondent au regroupement des individus selon la variable illustrative « site expérimental ».
L’application des digestats au sol soulève la question de leurs impacts sur le fonctionnement biologique des sols. Dans de nombreuses études, les effets des digestats sur le fonctionnement du sol ont été appréhendés à court terme dans les semaines post-épandage en conditions contrôlées ou in situ. Dans les semaines suivant l’application de digestats au sol, certains auteurs ont mis en évidence une augmentation de la taille et de l’activité de minéralisation des micro-organismes du sol suggérant une utilisation rapide du C et de N des digestats (Odlare et al., 2008) et potentiellement une immobilisation nette d’N d’autant plus élevée pour des digestats à faible ratio C/N (Alburquerque et al., 2012 ; de la Fuente et al., 2013). Dans des pots de cultures, Walsh et al. (2012) mettent en évidence que l’application de digestat stimule la communauté bactérienne au même titre que les engrais minéraux. Dans notre cas, les effets des digestats sur le fonctionnement microbien du sol sont appréhendées en sortie d’hiver 2019 soit plusieurs mois après les derniers épandages de fin 2018. Aucun effet des modalités de fertilisations n’est mis en évidence. Ces résultats sont cohérents avec d’autres études sur des essais longue durée pour lesquels l'application de digestats ne modifie pas significativement l’abondance de bactéries et de champignons du sol ni leur diversité (Coehlo et al., 2019, 2020).
Conclusion et perspectives
Deux années après la mise en place de ce dispositif expérimental européen sur cinq sites de prairie permanente, dans des contextes pédoclimatiques différents, le constat est fait que les rendements en biomasse sont significativement plus élevés sur les parcelles fertilisées par 230 unités d’azote sous forme de nitrate d’ammonium en comparaison des parcelles non fertilisées. Les rendements en biomasse des parcelles fertilisées (230 unités) par du lisier brut ou du digestat sont intermédiaires.
Malgré cette différence de biomasse, les valeurs fourragères et les teneurs en éléments minéraux des fourrages récoltés ne permettent pas de discriminer les modalités de fertilisation. Les valeurs fourragères et les teneurs en éléments minéraux discriminent davantage les différents sites ou encore le numéro d’ordre de la fauche dans l’année. Par ailleurs, après deux années d’expérimentation, il n’est pas possible non plus de distinguer d’effet des modalités de fertilisation sur le fonctionnement microbien des sols. Le dispositif étudié ici est assez unique dans le sens où il compare cinq modalités de fertilisation (dont du digestat de méthanisation, du lisier et de l’ammonitrate) sur cinq sites aux conditions pédoclimatiques différentes. Le changement de pratique de fertilisation ne permet pas, à nombre d’unités d’azote équivalent apporté, d’assurer la productivité des prairies permanentes, mais permettent en revanche de maintenir la qualité des fourrages récoltés. Deux ans après la mise en place des différents sites, les variables microbiennes suivies, bien que considérées comme des indicateurs de réponse aux changements de pratiques agricoles, ne permettent pas de mettre en évidence d’effets significatifs sur le fonctionnement microbien des sols. Un point d’étape à 5 ans sera réalisé sur ces mêmes dispositifs afin d’évaluer, à moyen terme, l’impact de ce changement de pratique sur l’évolution de la teneur en carbone des sols, les activités microbiennes de ces sols et la qualité des fourrages récoltés.
Références
Abbott, L. K., & Murphy, D. V. (Eds.). (2003). Soil biological fertility: a key to sustainable land use in agriculture. Springer Science & Business Media.
Alburquerque, J. A., de la Fuente, C., & Bernal, M. P. (2012). Chemical properties of anaerobic digestates affecting C and N dynamics in amended soils. Agriculture, ecosystems & environment, 160, 15-22.
Coelho, J. J., Hennessy, A., Casey, I., Bragança, C. R. S., Woodcock, T., & Kennedy, N. (2020). Biofertilisation with anaerobic digestates: A field study of effects on soil microbial abundance and diversity. Applied Soil Ecology, 147, 103403.
Coelho, J. J., Hennessy, A., Casey, I., Woodcock, T., & Kennedy, N. (2019). Responses of ryegrass, white clover, soil plant primary macronutrients and microbial abundance to application of anaerobic digestates, cattle slurry and inorganic N-fertiliser. Applied Soil Ecology, 144, 112-122.
de la Fuente, C., Alburquerque, J. A., Clemente, R., & Bernal, M. P. (2013). Soil C and N mineralisation and agricultural value of the products of an anaerobic digestion system. Biology and Fertility of Soils, 49(3), 313-322.
Guilayn, F., Jimenez, J., Martel, J. L., Rouez, M., Crest, M., & Patureau, D. (2019). First fertilizing-value typology of digestates: A decision-making tool for regulation. Waste management, 86, 67-79.
Mäder, P., Fliessbach, A., Dubois, D., Gunst, L., Fried, P., & Niggli, U. (2002). Soil fertility and biodiversity in organic farming. Science, 296(5573), 1694-1697.
Manlay, R. J., Feller, C., & Swift, M. J. (2007). Historical evolution of soil organic matter concepts and their relationships with the fertility and sustainability of cropping systems. Agriculture, Ecosystems & Environment, 119(3-4), 217-233.
Möller, K., & Müller, T. (2012). Effects of anaerobic digestion on digestate nutrient availability and crop growth: a review. Engineering in Life Sciences, 12(3), 242-257.
Nesheim, L., Boller, B. C., Lehmann, J., & Walther, U. (1990). The effects of nitrogen in cattle slurry and mineral fertilizer on nitrogen-fixation by white clover. Grass and Forage Science, 45(1), 91–9.
Odlare, M., Pell, M., & Svensson, K. (2008). Changes in soil chemical and microbiological properties during 4 years of application of various organic residues. Waste management, 28(7), 1246-1253.
Petitjean, C., Philibert, A., Manneville, V., Amiaud, B., Perrin, A. S., Charrier, X., et al. (2018). Systèmes de polyculture-élevage: quels effets des pratiques agricoles sur les teneurs en matières organiques et le fonctionnement microbien du sol? Fourrages, 236, 239-247
Sparling, G., Vojvodić-Vuković, M., & Schipper, L. A. (1998). Hot-water-soluble C as a simple measure of labile soil organic matter: the relationship with microbial biomass C. Soil Biology and Biochemistry, 30(10-11), 1469-1472.
Vance E.D., Brookes P.C., Jenkinson D.S. (1987). An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 19, 703-707.
Zhang, J. B., Zhu, T. B., Cai, Z. C., Qin, S. W., & Müller, C. (2012). Effects of long‐term repeated mineral and organic fertilizer applications on soil nitrogen transformations. European Journal of Soil Science, 63(1), 75-85.
Zhang, Q., Zhou, W., Liang, G., Sun, J., Wang, X., & He, P. (2015). Distribution of soil nutrients, extracellular enzyme activities and microbial communities across particle-size fractions in a long-term fertilizer experiment. Applied Soil Ecology, 94, 59-71.

Les articles sont publiés sous la licence Creative Commons (CC BY-NC-ND 2.0)
Pour la citation et la reproduction de cet article, mentionner obligatoirement le titre de l'article, le nom de tous les auteurs, la mention de sa publication dans la revue AES et de son URL, la date de publication.