

Mécanismes de régulation biologique des adventices par les couverts végétaux
Delphine MOREAU* 1, Guillaume ADEUX 1, Alicia ROUGE 1,2, Jean-Philippe GUILLEMIN 1, Stéphane CORDEAU 1
1 Agroécologie, INRAE, Institut Agro, Univ. Bourgogne, Univ. Bourgogne Franche-Comté, F-21000 Dijon, France
2 AgroParisTech, 75005 Paris, France
Résumé
La régulation biologique des adventices par les couverts végétaux cultivés peut impliquer différents mécanismes. La compétition pour les ressources est le mécanisme considéré comme prépondérant dans la régulation des adventices par les couverts végétaux. L’allélopathie pourrait également jouer un rôle mais ses effets au champ sont difficiles à démontrer car difficiles à dissocier des effets d’autres mécanismes. Les couverts végétaux peuvent également affecter les adventices en modifiant le microclimat à la surface du sol.
La régulation des adventices par les couverts végétaux peut également se faire par l’intervention d’organismes auxiliaires, comme des insectes responsables de prédation (carabes) ou des microorganismes pathogènes responsables de parasitisme (par ex. champignons responsables de fontes de semis). Cependant, on ne sait pas à ce jour si ces mécanismes peuvent avoir un impact significatif sur la banque de graines et la flore adventice exprimée dans les cultures suivantes.
Pour aller plus loin dans la quantification des effets relatifs de ces différents mécanismes et la compréhension de leurs déterminants, des recherches sont en cours, alliant approches expérimentales au champ, en conditions contrôlées et modélisation.
Mots-clés : compétition, allélopathie, modification du microclimat, prédation, parasitisme
Summary
Different mechanisms may be involved in weed biological regulation crop canopies. Competition for resources (light, water and minerals) is the mechanism considered to be preponderant in the regulation of weeds by crop canopies. Allelopathy could also play a role, but its effects in the field are difficult to demonstrate because difficult to dissociate from the effects of other mechanisms, in particular competition (for allelopathy generated by living plants) and immobilization of nitrogen (for residue-generated allelopathy). Crop canopies can also affect weeds by modifying the microclimate at the soil surface (temperature, light and humidity). The regulation of weeds by plant cover can also be done indirectly, i.e. via auxiliary organisms such as insects responsible for predation (beetles) or pathogenic microorganisms responsible for parasitism (eg. phytopathogenous fungi). While predation is the subject of a growing number of studies, few demonstrate that this mechanism is intense enough to have a significant impact on the seed bank and on the weed flora that will be expressed in the following crops. Few works have been carried out on the role of parasitism by phytopathogenic fungi for weed regulation. Studies combining experimental approaches in the field, under controlled conditions and modelling, are in progress to go further in quantifying the relative effects of these different mechanisms and understanding their determinants.
Introduction
Les systèmes de culture agroécologiques ont pour objectif de valoriser les services rendus par les interactions entre organismes dans la parcelle agricole afin de réduire le recours aux intrants chimiques. Cet article propose une vue d'ensemble des mécanismes impliqués dans la régulation biologique des adventices (avec un focus sur les plantes adventices non parasites) par les couverts végétaux cultivés, qu’il s’agisse de cultures de rente ou de service (ex : couverts d’interculture).

Les mécanismes de régulation biologique des adventices par les couverts végétaux se répartissent en deux catégories (Figure 1) :
- Les mécanismes à effet direct regroupent la compétition pour les ressources (i.e. lumière et ressources du sol), l’allélopathie (libération de composés biochimiques par une plante vivante ou par les résidus de la plante après sa destruction), et la modification du microclimat (e.g. lumière, température, humidité) à la surface du sol qui, sans pour autant exercer de compétition, impacte les adventices.
- Les mécanismes à effet indirect faisant intervenir des organismes auxiliaires, comme des insectes responsables de prédation (carabes) ou des microorganismes pathogènes responsables de parasitisme (par ex. champignons responsables de fontes de semis).
Compétition
La compétition pour les ressources (lumière, eau, nutriments) est le mécanisme considéré comme prépondérant dans la régulation des adventices par les couverts végétaux. Elle se produit lorsque les plantes cultivées et adventices partagent un pool de ressources commun, dans l'espace et dans le temps, et qui est insuffisant pour répondre aux besoins de toutes les plantes (Zimdahl, 2004).
La lumière a longtemps été considérée comme la principale ressource pour laquelle la compétition s'exerce dans les systèmes intensifs sous climat tempéré. Cependant, la compétition pour les ressources du sol peut être très importante dans les couverts d’interculture ; en effet ils sont souvent implantés en été, période pendant laquelle les ressources en nutriments (i.e. azote) et en eau peuvent être faibles. De manière plus générale, la compétition pour l’eau et l’azote pourrait jouer un rôle accru dans les parcelles agricoles dans le contexte de changement climatique et de réduction des intrants chimiques (engrais minéraux).
La compétition générée par les couverts cultivés sur les adventices dépend :
- Des dates de levée relatives des différentes plantes composant la communauté (Knezevic et al., 2017). Les adventices les plus nuisibles sont généralement celles qui lèvent au même moment que le couvert cultivé. Celles qui lèvent avant étant le plus souvent détruites lors d’opérations culturales, alors que celles qui lèvent après subissent la compétition générée par le couvert déjà en place.
- Du niveau de disponibilité en ressources, qu’il s’agisse de la lumière (pour une plante donnée, l’accès à la lumière dépend fortement de son positionnement hiérarchique dans le couvert), de l’eau et des minéraux.
- De la dynamique des besoins des plantes (Lemaire et al., 2007). Ces besoins sont dépendants de la vitesse et du potentiel de croissance des plantes. Par exemple, une culture avec un fort potentiel de croissance, en particulier une forte vitesse de croissance en début de cycle, aura une forte vitesse de captation des ressources et donc un pouvoir compétitif accru.
- De la capacité des plantes à prélever les ressources. Ainsi, une colonisation du sol rapide en largeur et en profondeur, avec une forte densité de racines et/ou une forte capacité d’absorption par les racines seront favorables à un accroissement du pouvoir compétitif (Hodge et al., 2009).
- De la capacité des plantes à s'adapter à une limitation des ressources, en modifiant par exemple leur morphologie et/ou l’allocation de biomasse entre les différents compartiments (Brouwer, 1962 ; Colbach et al., 2020 ; Perthame et al., 2022).
L’intensité et l’issue de la compétition sont multifactorielles, dépendant des espèces/variétés, des itinéraires techniques appliqués, et plus généralement des systèmes de culture. Le contexte pédoclimatique et floristique interfère également. Pour appréhender cette complexité, le modèle mécaniste de simulation FLORSYS a été développé (Colbach et al., 2021). Il simule les effets des systèmes de culture en interaction avec le pédoclimat sur la dynamique des adventices et leur impact en particulier sur le rendement des cultures. Il s'agit d'une parcelle expérimentale virtuelle sur laquelle l'utilisateur peut tester de nombreux systèmes de culture sur plusieurs années, avec différents scénarios (météo, sol, flore), pour évaluer la performance en termes de production agricole, de biodiversité et nuisibilité liée aux adventices. Pour ce faire, le modèle simule au jour le jour et en 3D les processus du cycle de vie individuellement pour chaque plante de la parcelle (adventices et cultivées) sur plusieurs années/décennies, à partir des opérations culturales, du pédoclimat et du stock de semences adventices dans le sol. Des simulations dans différentes situations (zones géographiques et systèmes de culture différents) sans limitation en eau et en azote ont été réalisées (Colbach et al., 2019). Il ressort qu’en général les cultures les plus efficaces pour réprimer les adventices par compétition pour la lumière sont notamment celles qui s’installent rapidement avec des feuilles fines et larges (plutôt que petites et épaisses), qui ont tendance à croître en largeur et qui sont capables d'adapter leur morphologie en cas d'ombrage par les plantes voisines. A noter que ces tendances générales obtenues sur un grand nombre de situations ne sont pas forcément adaptées aux spécificités de chacune des situations de production.
Le rôle de la compétition pour l’azote dans la régulation des adventices par les couverts cultivés est moins bien connu. Une analyse détaillée de la littérature scientifique (Perthame, 2020) indique que, dans la moitié des articles analysant les effets de la fertilisation azotée sur les rapports culture-adventices, une réduction de la quantité d’azote apportée favorise les adventices au détriment des cultures. Un quart des études montre l’inverse alors qu’aucun effet n’est observé pour le dernier quart. Il est probable que ces différences proviennent de différences entre les situations d’études (adventices, espèces/variétés cultivées, systèmes de culture, conditions pédoclimatiques).
Afin de mieux comprendre les relations de compétition pour l’azote, des travaux ont été menés (Moreau et al., 2013 ; Moreau et al., 2014). Les espèces adventices les plus problématiques en culture sont généralement des espèces très nitrophiles (Fried, 2007), car elles occupent préférentiellement des habitats riches en azote. Ces travaux visaient à comprendre le succès des espèces nitrophiles dans les parcelles cultivées (généralement riches en azote). Ils montrent que, plus une espèce est nitrophile, plus son potentiel de croissance aérienne est élevé et plus sa croissance dépend de la disponibilité en azote du sol. En outre, moins elle alloue de biomasse à ses racines (par rapport à la partie aérienne) mais chaque unité de racine est plus efficiente à prélever l’azote du sol. Ainsi, la forte capacité des espèces nitrophiles à prélever l’azote en situation de forte disponibilité en azote, permet une expansion rapide de leur surface foliaire. Cela leur confère (1) une forte interception de lumière et donc une forte capacité de photosynthèse, et (2) une forte capacité d’ombrage vis-à-vis des plantes voisines moins nitrophiles. Elles apparaissent donc comme de très bonnes compétitrices pour la lumière.
Allélopathie
L’allélopathie est un mécanisme par lequel une plante (vivante ou ses résidus en décomposition) affecte la croissance des plantes voisines par la production de composés chimiques libérés dans l'environnement (Inderjit and Duke, 2003). Alors que de nombreuses études montrent des effets allélopathiques en laboratoire, les effets au champ sont difficiles à démontrer. Les effets de l’allopathie générée par les résidus de culture sont difficiles à dissocier des effets relatifs à l’immobilisation de l’azote par les résidus de culture (Doré, 2004). En ce qui concerne l’allélopathie par les plantes vivantes, la difficulté est de dissocier les effets de l’allélopathie de ceux de la compétition pour les ressources. En effet, une proximité des racines des plantes cultivées et adventices est nécessaire pour que des effets allélopathiques soient décelables. Or une telle situation favorise la compétition pour les ressources.
Ainsi, la première revue systématique de la littérature scientifique a été menée pour objectiver l’existence d’éléments de preuve au champ des effets de l’allélopathie dans la régulation des adventices, indépendamment de la compétition (Mahé et al., 2022). Pour se prémunir le plus possible des effets de la compétition, la recherche bibliographique s’est focalisée sur des études au champ comparant différentes variétés au sein d’une même espèce cultivée. En effet, des variétés peuvent être choisies pour avoir des propriétés allélopathiques différentes (préalablement caractérisées en conditions contrôlées). Des effets de l’allélopathie pourront être attestés si les différences de régulation des adventices observées entre variétés au champ sont corrélées aux différences de propriétés allélopathiques, sans corrélation avec des traits des variétés relatifs à la compétition.
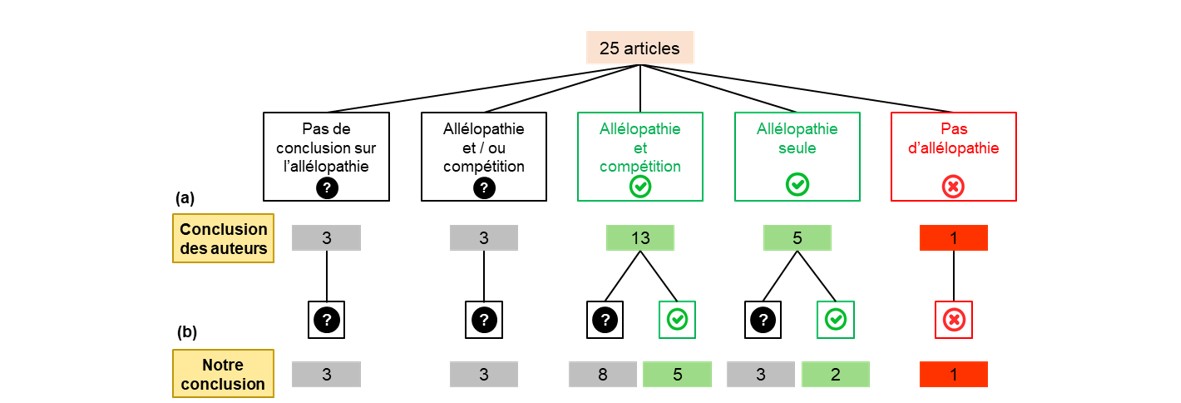
Notre analyse montre qu’une grande partie des articles recensés ignore le mécanisme de compétition, attribuant de manière non objective l’ensemble des effets observés à l’allélopathie. Seuls 25 articles (sur 42 initialement) considèrent explicitement allélopathie et compétition (Figure 2). Pour 13 d’entre eux, les auteurs ont conclu qu’à la fois l’allélopathie et la compétition expliquaient les différences de régulation des adventices entre variétés. Dans cinq autres, l’allélopathie a été considérée comme le seul mécanisme à l’œuvre. Dans un dernier article, les différences entre variétés ont été attribuées à la compétition seule. Une analyse détaillée de ces articles a été menée par une approche qualitative (pertinence des dispositifs et protocoles expérimentaux) et quantitative (gammes de variation des variables mesurées, analyses de corrélation). Nous avons identifié que, parmi les articles rapportant un effet de l’allélopathie (seule ou combinée à la compétition), seuls sept pouvaient fournir des éléments de preuve des effets de l’allélopathie au champ. Nous avons néanmoins émis quelques réserves liées principalement à l’absence de mesures permettant de s’assurer que les ressources du sol n’étaient pas limitantes (condition importante pour s’assurer de l’absence de compétition pour les ressources du sol). Pour aller plus loin, il sera nécessaire de mettre en place de nouvelles expérimentations mobilisant les recommandations de Mahé et al. (2022) en matière de dispositif expérimental, variables à mesurer et analyse statistique des données.

Modification du microclimat à la surface du sol
Les couverts végétaux sont en capacité de modifier le microclimat à la surface du sol, notamment en termes de température, lumière et humidité. Ces facteurs environnementaux influençant notamment la germination et l’émergence des adventices, les couverts végétaux peuvent jouer un rôle dans la régulation des adventices. Environ la moitié des espèces adventices sont photosensibles, c’est-à-dire nécessitent de la lumière pour germer. La présence d’un couvert végétal peut réduire l’exposition à la lumière, et également modifier sa qualité (diminution du ratio rouge clair/rouge sombre), ce qui peut affecter les dynamiques de germination et d’émergence. Ainsi, des travaux ont montré qu’en présence d’un couvert végétal, l'émergence des adventices a tendance à diminuer de manière significative (-17,1% en comparaison d’un témoin sans couvert) (Figure 3) mais de manière variable entre les espèces adventices (Cordeau et al., 2015 ; Cordeau et al., 2018).
Prédation
La prédation est une interaction qui relève d’une approche verticale entre niveaux trophiques : l’herbivorie et plus spécifiquement la granivorie par des organismes consommateurs de graines. La flore adventice est composée essentiellement d’espèces annuelles produisant d’importantes quantités de graines tombant au sol qui réalimentent le stock semencier chaque année. Ces graines représentent une ressource alimentaire importante dans les agroécosystèmes et de fait, de nombreux arthropodes tels les coléoptères carabiques (Figure 4), criquets, fourmis ainsi que des petits mammifères et oiseaux consomment des quantités non négligeables de graines d’adventices.
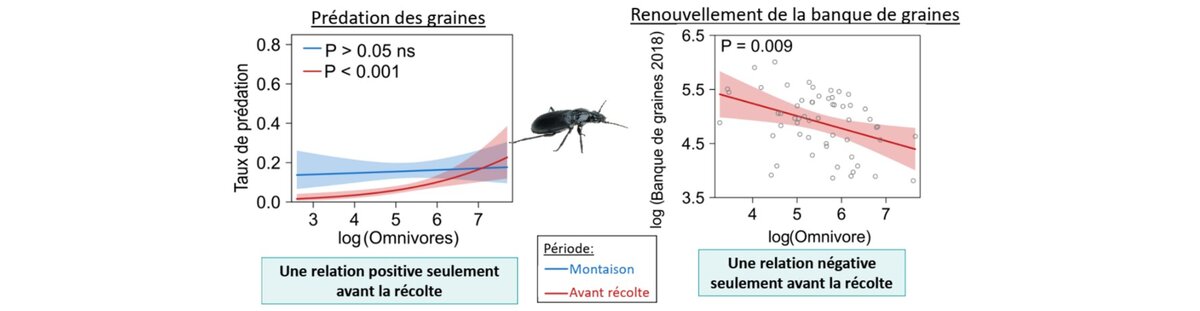
Depuis quelques années, des équipes de recherche étudient les déterminants de la prédation des graines d’adventices par des organismes granivores ou omnivores naturellement présents dans les champs cultivés (Kulkarni et al., 2015). Sur la base d’une dizaine d’études menées en Europe et aux Etats-Unis qui fournissent des mesures de prédation répétées en parcelles agricoles, le taux annuel de prédation (incluant des prédateurs vertébrés et invertébrés) est d’environ 40% mais il est assez variable en fonction des situations de parcelles, ici de 8 à 70% en fonction des espèces adventices et du contexte agronomique (Davis et al., 2011). Des travaux de modélisation sur la dynamique des communautés adventices suggèrent qu’une perte annuelle de 25 à 50% des graines adventices serait suffisante pour affecter la dynamique des populations et les faire régresser.
Dans les régions tempérées, les carabidés (Adephaga, Carabidae) sont souvent les principaux invertébrés consommateurs de graines d’adventices (Honek et al., 2003) et la plupart des études portent sur la relation graines d’adventices/carabes. La présence d'un couvert végétal, qu’il s’agisse de couverts installés en interculture (Davis and Liebman, 2003) ou de couverts permanents (Blubaugh et al., 2016), peut favoriser des insectes auxiliaires notamment les carabes en leur fournissant un abri et des ressources. Même s’il en existe, encore peu de travaux démontrent que cette régulation est suffisamment intense pour avoir un impact significatif sur la banque de graines et sur la flore adventice qui va s’exprimer dans les cultures suivantes.

Parasitisme
La présence d’un couvert végétal peut induire une augmentation de l’humidité de l’air et, selon les périodes, une diminution de l’humidité du sol. Un couvert peut aussi tamponner la température au sol, et en particulier diminuer sa température en période chaude en limitant l’intensité du rayonnement solaire atteignant le sol. Ces conditions environnementales générées par la présence de couverts végétaux sont a priori plus favorables aux champignons qui agissent sur les adventices de différentes manières (Figure 5). Parmi ces champignons, certains sont phytopathogènes des semences et/ou des plantes adventices (Hatcher and Melander, 2003). Très peu de données sont disponibles sur cet aspect et peu de travaux sont menés sur ce sujet (Triolet et al., 2019). En effet les phytopathologistes ont longtemps concentré leur travail sur des espèces végétales cultivées. L’accroissement des connaissances sur l’action de champignons phytopathogènes sur les adventices pourrait permettre de mieux quantifier les effets de ce mécanisme et ses déterminants, afin de déterminer s’il pourrait jouer un rôle pour favoriser la régulation des adventices sans affecter les cultures de rente.
Conclusion
En conclusion, la compétition est souvent considérée comme un mécanisme majeur dans la régulation des adventices, notamment la compétition pour la lumière. L’allélopathie, la prédation et le parasitisme jouent vraisemblablement aussi un rôle mais leurs effets relatifs ont à ce jour été peu quantifiés et leurs déterminants restent à préciser. Des travaux alliant approches expérimentales au champ, en conditions contrôlées et modélisation, sont en cours pour aller plus loin dans cette direction.
Références bibliographiques
Blubaugh, C. K., Hagler, J. R., Machtley, S. A., Kaplan, I., 2016. Cover crops increase foraging activity of omnivorous predators in seed patches and facilitate weed biological control. Agriculture, Ecosystems & Environment 231, 264-270.
Brouwer, R., 1962. Nutritive influences on the distribution of dry matter in the plant. 10, 399-408.
Carbonne, B., 2020. Le rôle des interactions biotiques dans un système proie-prédateur: le cas de la prédation et de la régulation des graines d’adventices par les carabes. Université Bourgogne Franche-Comté http://www.theses.fr/2020UBFCK047/document,
Colbach, N., Gardarin, A., Moreau, D., 2019. The response of weed and crop species to shading: Which parameters explain weed impacts on crop production? Field Crops Research 238, 45-55.
Colbach, N., Moreau, D., Dugué, F., Gardarin, A., Strbik, F., Munier-Jolain, N., 2020. The response of weed and crop species to shading. How to predict their morphology and plasticity from species traits and ecological indexes? European Journal of Agronomy 121, 126158.
Colbach, N., Colas, F., Cordeau, S., Maillot, T., Queyrel, W., Villerd, J., Moreau, D., 2021. The FLORSYS crop-weed canopy model, a tool to investigate and promote agroecological weed management. Field Crops Research 261, 108006.
Cordeau, S., Guillemin, J. P., Reibel, C., Chauvel, B., 2015. Weed species differ in their ability to emerge in no-till systems that include cover crops. Annals of Applied Biology 166, 444–455.
Cordeau, S., Wayman, S., Reibel, C., Strbik, F., Chauvel, B., Guillemin, J. P., 2018. Effects of drought on weed emergence and growth vary with seed burial depth and presence of a cover crop. Weed Biology and Management 18, 12-25.
Davis, A. S., Liebman, M., 2003. Cropping system effects on giant foxtail (Setaria faberi) demography: I. Green manure and tillage timing. Weed Science 51, 919-929.
Davis, A. S., Daedlow, D., Schutte, B. J., Westerman, P. R., 2011. Temporal scaling of episodic point estimates of seed predation to long‐term predation rates. Methods in Ecology and Evolution 2, 682-890.
Doré, T. S. M. P. F. G. C., 2004. Approche agronomique de l'allélopathie. Cahiers Agricultures 13, 249-256.
Fried, G., 2007. Variations spatiales et temporelles des communautés adventices des cultures annuelles en France. Thèse de l'université de Bourgogne, Dijon.
Hatcher, P., Melander, B., 2003. Combining physical, cultural and biological methods: prospects for integrated non‐chemical weed management strategies. Weed Research 43, 303-322.
Hodge, A., Berta, G., Doussan, C., Merchan, F., Crespi, M., 2009. Plant root growth, architecture and function. Plant and Soil 321, 153-187.
Honek, A., Martinkova, Z., Jarosik, V., 2003. Ground beetles (Carabidae) as seed predators. European Journal of Entomology 100, 531-544.
Inderjit, Duke, S. O., 2003. Ecophysiological aspects of allelopathy. Planta 217, 529-539.
Knezevic, S. Z., Evans, S. P., Blankenship, E. E., Van Acker, R. C., Lindquist, J. L., 2017. Critical period for weed control: the concept and data analysis. Weed Science 50, 773-786.
Kulkarni, S. S., Dosdall, L. M., Willenborg, C. J., 2015. The role of ground beetles (Coleoptera: Carabidae) in weed seed consumption: a review. Weed Science 63, 335-376.
Lemaire, G., van Oosterom, E., Sheehy, J., Jeuffroy, M. H., Massignam, A., Rossato, L., 2007. Is crop N demand more closely related to dry matter accumulation or leaf area expansion during vegetative growth? Field Crops Research 100, 91-106.
Mahé, I., Chauvel, B., Colbach, N., Cordeau, S., Gfeller, A., Reiss, A., Moreau, D., 2022. Deciphering field-based evidence for weed regulation by crop allelopathy. A review. Agronomy for Sustainable Development 42, 50.
Moreau, D., Milard, G., Munier-Jolain, N., 2013. A plant nitrophily index based on plant leaf area response to soil nitrogen availability. Agronomy for Sustainable Development 33, 809-815.
Moreau, D., Busset, H., Matejicek, A., Munier-Jolain, N., 2014. The ecophysiological determinants of nitrophily in annual weed species. Weed Research 54, 335-346.
Perthame, L., 2020. Analyse et modélisation du rôle de la compétition pour l’azote dans la régulation des adventices. Université Bourgogne Franche-Comté http://www.theses.fr/2020UBFCK049/document,
Perthame, L., Colbach, N., Busset, H., Matejicek, A., Moreau, D., 2022. Morphological response of weed and crop species to nitrogen stress in interaction with shading. Weed Research 62, 160-171.
Triolet, M., Guillemin, J.-P., Andre, O., Steinberg, C., 2019. Fungal-based bioherbicides for weed control: a myth or a reality? Weed Research
Zimdahl, R. L., 2004. The Elements of Competition. In: Weed-Crop Competition, Blackwell Publishing Professional,

Les articles sont publiés sous la licence Creative Commons (CC BY-NC-ND 2.0)
Pour la citation et la reproduction de cet article, mentionner obligatoirement le titre de l'article, le nom de tous les auteurs, la mention de sa publication dans la revue AES et de son URL, la date de publication.