

De l’intérêt des approches comparatives en agronomie
The interest of comparative approaches in agronomy
Stark, F.1, Kebede Y.2, Poeydebat, C.3, Doré, T.4
1 SELMET, Univ Montpellier, INRAE, CIRAD, L’Institut Agro – Montpellier SupAgro, Montpellier, France
2 Eco&Sol, Univ Montpellier, INRAE, CIRAD, IRD, L’Institut Agro – Montpellier SupAgro, Montpellier, France
3 UPR GECO, CIRAD, F-97285 Le Lamentin, Martinique, France
4 Université Paris-Saclay, AgroParisTech, INRAE, UMR Agronomie, 78850, Thiverval- Grignon, France.
Auteur correspondant : fabien.stark@inrae.fr
Résumé
Les sciences agronomiques ont évolué au cours du temps et se sont enrichies au gré des échelles d’analyse, des situations et des disciplines connexes qui ont permis d’en élargir la portée. L’approche comparative en agronomie, objet du présent article, est l’un des développements de la discipline agronomique qu’il nous semble pertinent d’approfondir. C’est une approche qui cherche à tirer parti de la diversité du fonctionnement des agroécosystèmes, et qui pourrait être utilisée pour explorer les processus agronomiques liant structures, fonctionnement, propriétés et performances des agroécosystèmes. Il s’agirait de mettre en regard des situations variées pour identifier ce qui, dans la structure et le fonctionnement des agroécosystèmes, détermine leurs propriétés, et influe sur leurs performances. Nous proposons pour ce faire de remobiliser trois exemples d’études comparatives récentes en agronomie qui, bien que n’ayant pas été explicitement conçues dans cet objectif, permettent d’en discuter la portée et de proposer des pistes pour de futurs travaux. Ces trois exemples concernent des travaux réalisés dans différents contextes socio-écologiques (Amérique du Sud-Caraïbe-Afrique de l’Est) et renvoient à des échelles d’analyse différentes (parcelle-exploitation-paysage). Les trois études de cas présentées dans cet article montrent certaines similitudes dans les démarches mises en œuvre. En effet, ces travaux s’appuient sur des gradients de situations socio-écologiques pour comparer des processus (intégration agriculture élevage, diversité végétale, configuration paysagère), s’exprimant de manière différenciée, en fonction de ces situations. Dans tous les cas, la complexité des processus étudiés, s’inscrivant dans des situations réelles aux interactions multiples, nécessite d’avoir recours à des proxys pour répondre aux questions de recherche traitées. Dans une vision dynamique des activités de recherche et de développement, ces travaux permettent de faire émerger des propriétés agroécologiques pertinentes qu’il reste ensuite nécessaire d’approfondir pour monter en généricité.
Mots clés : gradient ; systémique ; comparaison ; agriculture ; agroécologie
Abstract
The science of agronomy has evolved over time and have has been enriched by analysis on various spatial scales and related disciplines which have made it possible to broaden its scope. The comparative approach in agronomy, the subject of this article, is one of the developments in the agronomy that we feel needs further attention. It is an approach that seeks to take advantage of the diversity of agroecosystem functioning and could be used to explore agronomic processes linking the structures, functioning, properties and performance of agroecosystems. It would involve comparing various situations to identify what, in the structure and functioning of agroecosystems, determines their properties and influences their performance. To this end, we propose to remobilize three examples of recent comparative studies in agronomy which, although not explicitly designed with this objective in mind, allow us to discuss their scope and propose avenues for future work. . These three examples concern work carried out in different socio-ecological contexts (South America-Caribbean-East Africa) and refer to different scales of analysis (plot-farm-landscape). The three case studies presented in this article show certain similarities in the approaches implemented. Indeed, this work is based on gradients of socio-ecological situations comparing processes (integration of agriculture and livestock farming, plant diversity, landscape configuration), which are expressed in a differentiated manner depending on these situations. In all cases, the complexity of the processes studied, which take place in real situations with multiple interactions, requires the use of proxies to answer the research questions addressed. In a dynamic vision of research and development activities, this work allows relevant agroecological properties to emerge, which then need to be studied in greater depth in order to become more generalized.
Key words: gradient; systemic; comparison; agriculture; agroecology
Introduction
Les agronomes ont un double intérêt pour la relation de l’agriculture à l’espace. D’une part, ils étudient et participent au pilotage d’objets correspondant à des entités spatiales délimitées de manières différentes (en particulier parcelle, exploitation agricole, différents types de territoires), et dont les acteurs diffèrent. Indépendamment de l’échelle, car chacune de ces entités peut correspondre à des aires extrêmement variées, ce qui est au cœur du travail des agronomes c’est la manière dont, dans chacune d’entre elles, l’action de l’homme interagit avec les caractéristiques physiques et biologiques de l’agroécosystème. D’autre part, les agronomes cherchent à comparer les systèmes mis en œuvre dans différentes régions du monde, différents milieux, différentes sociétés, en visant l’enrichissement de la théorie agronomique par sa mise à l’épreuve dans une diversité de situations, pour envisager l’extrapolation de pratiques d’un endroit à un autre (voir un exemple récent dans Cann et al. (2020), pour une adaptation des pratiques agricoles aux changements climatiques en substituant le temps par l’espace, Holzkämper et al., 2011) - et également par curiosité envers la diversité des agricultures.
Le présent article envisage une troisième posture, qui cherche à tirer parti de la diversité du fonctionnement des agroécosystèmes pour élaborer des règles générales de fonctionnement liant structures, propriétés et performances des systèmes, suivant ce que Doré (2010) et Doré et al. (2011) ont proposé d’appeler une « agronomie comparative ». Il s’agirait de mettre en regard des situations variées pour déterminer ce qui, dans la structure des agroécosystèmes, détermine leurs propriétés, et influe sur leurs performances. A notre connaissance, il n’existe pas d’approche formalisée visant ces objectifs. Sans donner de définition formelle à ces termes, manipulés différemment par les scientifiques, Doré (2010) et Doré et al. (2011) qualifient de « performances » la façon dont ces systèmes, dans une gamme de situations naturelles (notamment climatiques) et socio-économiques (notamment de marché), satisfont un certain nombre de services : production alimentaire et non alimentaire, bienfaits environnementaux et écologiques, etc. Ils considèrent que la recherche de performances suppose de s’appuyer, comme le propose Bonaudo et al. (2014), sur des propriétés émergentes issues de processus écologiques des agrosystèmes, en particulier de leur potentiel de productivité, de leur autonomie (pour limiter les externalités environnementales négatives), de leur résilience (pour faire face à une diversité d’aléas mais aussi pour mieux s’adapter au changement permanent). Sur cette base, deux hypothèses majeures nous semblent étayer la suggestion faite par ces auteurs de progresser dans la connaissance des relations structure-propriétés-performances des agroécosystèmes.
La première est qu’améliorer ces propriétés passe non seulement par une modification des choix techniques appréhendés à différentes échelles spatiales et temporelles, mais aussi plus profondément par une transformation de certaines caractéristiques structurelles des agroécosystèmes comme leur diversité biologique, l’organisation de cette diversité dans l’espace et le temps, ou encore le rythme et l’intensité des perturbations induites par les pratiques agricoles et d’élevage. L’identification des relations de causalité entre ces modes de conduite et les propriétés reste complexe à établir, compte tenu de la nature systémique et holistique des processus à l’œuvre. À titre d’exemple, les interactions biotiques au sein des agroécosystèmes sont complexes ; en obtenir une compréhension complète et intégrative est essentiel mais cela reste difficile à atteindre via des approches expérimentales classiques. La seconde hypothèse est qu’une approche comparative d’agroécosystèmes peut permettre de réaliser un saut qualitatif significatif dans la mise à jour et l’explicitation des relations entre structures et propriétés des agroécosystèmes – ce qui a été le cas dans d’autres disciplines (par exemple en biologie végétale et écophysologie (Berger et al., 2002 ; Mauseth, 2006) ; en sciences de l’évolution (Zink, 2002 ; Schluessel et al., 2008), en écologie marine (Thomson & Simanek, 1977 ; Fuhrman & Steele, 2008) ; en anatomie (Rosencrans et al., 2018).
En agronomie, il s’agirait de mener des comparaisons d’agroécosystèmes balayant une gamme large de conditions écologiques (notamment climats, ressources disponibles), en s’inspirant d’un procédé de macro-analyse largement utilisé dans d’autres disciplines, en particulier en écologie (voir des exemples dans Garnier et al., 2007 ; Huey et al., 2019). De même qu’il serait trop long de tester expérimentalement des hypothèses sur des processus évolutifs expliquant la diversité anatomique et/ou biologique, il serait long et coûteux de mettre en place des systèmes de production expérimentaux (d’autant qu’on ne sait pas forcément a priori quels éléments des systèmes faire varier) pour étudier leur fonctionnement. Il est plus avantageux d’utiliser et de juxtaposer des systèmes de production existants, comparables mais contrastés sur certains points. Cela se fait par exemple en foresterie (qui implique des systèmes de production au cycle pourtant plus long) avec des sites expérimentaux tels que ORPHEE[1] dans le sud-ouest de la France, qui est conçu pour étudier et comprendre l’effet de la diversité des forêts sur leur fonctionnement. Le réseau TreeDivNet[2] met en relation une série de sites expérimentaux de ce genre dans un but comparatif et de compréhension des liens de causalité entre configuration et gestion des forêts et fonctionnement des forêts.
Dans la suite de l’article sont exposés trois exemples d’études mobilisant des comparaisons de systèmes au cours de la dernière décennie (tableau 1), puis une discussion tire les enseignements synthétiques issus de ces exemples, en proposant des voies pour de futurs travaux, qui pourraient déboucher sur une réelle démarche formalisée d’agronomie comparative.
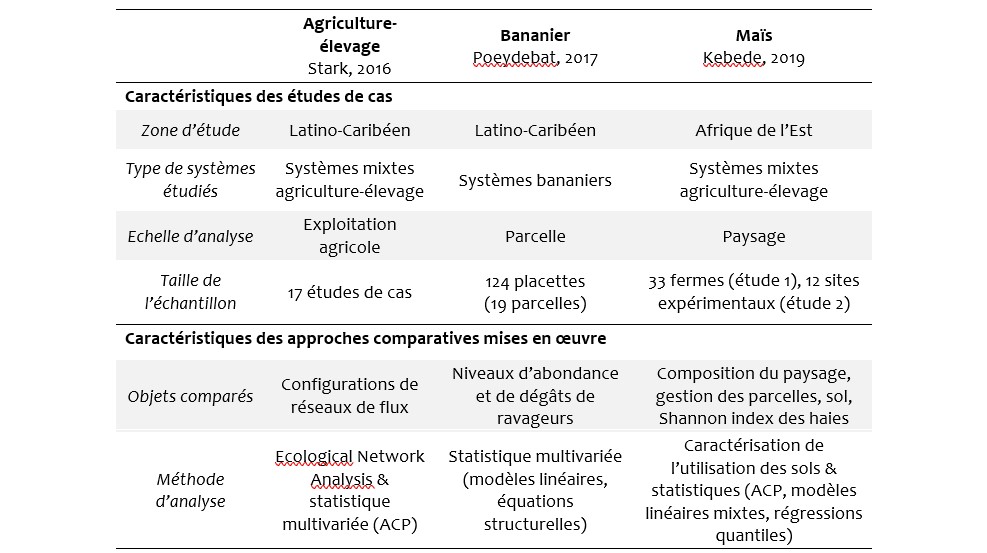
Tableau1 : Caractéristiques des trois études de cas
Analyse de l’effet de la structure de systèmes polyculture-élevage en milieu tropical sur leurs propriétés agroécologiques : le rôle des flux d’azote
Objectif et problématique
Pour s’adapter à la rareté croissante des ressources naturelles tout en répondant aux demandes d'une population mondiale en expansion, les systèmes agricoles doivent produire plus et mieux. Pour cela, la conception de systèmes agricoles locaux durables est primordiale. Ces systèmes agricoles devraient être à la fois efficaces, productifs, résilients et autosuffisants, sur la base des principes de l'agroécologie (Bonaudo et al., 2014). Au niveau d’un système agricole, l’intégration agriculture-élevage, en tant qu'ensemble de pratiques agricoles mobilisant des processus écologiques, peut contribuer à la performance agroécologique des systèmes mixtes agriculture-élevage, et au-delà, aux enjeux agricoles dans leur ensemble (González-García et al., 2012). La présente étude de cas avait pour objectif d’identifier les propriétés émergentes de systèmes agricoles complexes induites par les pratiques d’intégration agriculture-élevage, en particulier de savoir si ces pratiques permettent aux systèmes mixtes d’être agro-écologiquement performants (Stark et al., 2019).
L’hypothèse sous-jacente est que des systèmes agricoles diversifiés et intégrés mobilisent des processus biologiques et écologiques qui leurs confèrent des propriétés dites agroécologiques d’intérêt. En particulier, nous avons fait l’hypothèse :
- que l’autonomie serait liée au niveau d’intégration (par substitution d’intrants externes par le recyclage interne des ressources) ;
- d’un niveau d’équilibre efficience/résilience, sorte d’optimum de fonctionnement des systèmes agricoles. En deçà de cet équilibre, des situations pour lesquelles les systèmes étaient efficients au détriment de la résilience ; au-delà, des situations pour lesquelles les systèmes étaient résilients au détriment de l’efficience.
Dispositif de recherche
Comparer des systèmes agricoles présentant des structures (productions animales et végétales) et des fonctionnements (pratiques d’intégration agriculture-élevage) différents, implique de définir sur quelle base la comparaison peut se faire. Dans notre cas, la traduction des interactions agriculture-élevage en réseaux de flux de nutriment (ici, l’azote) en rend l’analyse et la comparaison possible et pertinente (Rufino et al., 2009). À travers l’Ecological Network Analysis (ENA), il est en effet possible de modéliser chaque système étudié en matrice de flux, de calculer un ensemble d’indicateurs caractérisant ce réseau de flux (intensité et organisation) et d’estimer les propriétés émergentes induites : résilience, efficience, autonomie et productivité (Ulanowicz et al., 2009). Dix-sept études de cas issues de trois territoires contrastés (Guadeloupe, Amazonie brésilienne et Cuba) représentant une gamme de réseaux de flux aux caractéristiques contrastés, ont été analysées et comparées. Cela a permis de préciser la structure, le fonctionnement et les performances des systèmes ainsi étudiés.
Résultats marquants
Cette mise en comparaison nous a tout d’abord permis de rendre compte de réseaux de flux d’azote aux caractéristiques contrastées, et de relier ces caractéristiques à la structure et au fonctionnement des systèmes mixtes agriculture-élevage étudiés :
- organisation du réseau de flux : connectivité entre les composantes du système et répartition de l’azote au sein de ce réseau de flux ;
- intensité du réseau de flux : part des flux d’azote qui circulent entre les compartiments (intensité des pratiques d’intégration agriculture-élevage par rapport aux flux totaux).

Figure 1 : Quatre configurations « type » des réseaux de flux d’azote en fonction du niveau d’intensité de l’azote circulant entre les compartiments et en fonction de l’organisation du réseau de flux (hétérogène/homogène). Source : Stark, 2016
La combinaison de ces deux caractéristiques a permis de dresser une typologie des configurations de réseaux de flux contrastés (Figure 1), allant de systèmes mixtes peu connectés (G5 en Guadeloupe) à des situations fortement connectées au sein desquelles les flux d’azote sont conséquents (C5 à Cuba), en passant par des situations très connectées pour lesquels les flux sont par ailleurs de faible intensité (B2 en Amazonie brésilienne). De plus, ces résultats montrent que les parts que prennent respectivement les productions animales et les productions végétales dans le système mixte jouent un rôle central dans les échanges de flux possibles, centralisant certains échanges (cas ensilage par exemple) ou au contraire favorisant une répartition homogène des flux entre productions (cas alimentation à partir de résidus de cultures par exemple).
Enfin, l’analyse (par analyse en composante principale) des corrélations entre les caractéristiques des réseaux de flux et les propriétés agroécologiques, en l’occurrence l’efficience, la résilience, la productivité et l’autonomie des systèmes étudiés, a permis de dégager différents profils de systèmes (Figure 2) :
- Des systèmes à haut niveau d’intrant, peu résilients et peu efficients malgré de bonnes productivités ; ils mettent peu de pratiques d’intégration de faible intensité en œuvre. Il s’agit de la majorité des exploitations guadeloupéennes ;
- Des systèmes à bas niveaux d’intrants et à bas niveau de productivité, mais résilients. Ces systèmes mettent en œuvre toute une diversité de pratiques d’intégration, mais de faible intensité. Il s’agit d’exploitations brésiliennes et de deux exploitations cubaines ;
- Des systèmes qui sont efficients, productifs grâce aux intrants et à l’intensité de l’intégration, avec un niveau de résilience intermédiaire. Ils mettent en œuvre une diversité de pratiques d’intégration d’intensité conséquente. Il s’agit de deux exploitations cubaines et d’une exploitation guadeloupéenne.
Principales conclusions
La comparaison de réseaux de flux de nutriments a permis l’analyse d’une gamme de situations contrastées, suivant plusieurs dimensions : intensité des flux, organisation des flux, accès aux facteurs de production et diversité des productions.
Sur cette base, il a ainsi été possible d’identifier certaines relations entre structure, fonctionnement et performances attendues de ces systèmes mixtes :
- La résilience des systèmes étudiés est fortement liée à l’organisation du réseau de flux, suivant deux dimensions. Tout d’abord, « l’équilibre » du réseau de flux, à savoir une répartition homogène de l’azote entre les flux, favoriserait la résilience du système, illustration de l’adage « ne pas mettre tous ses œufs dans le même panier ». Ensuite, le niveau de connectivité entre les compartiments du système et la redondance des « chemins » conféreraient au système la capacité à s’adapter en cas d’aléas sur l’un de ces flux.
- La productivité du système est quant à elle liée à l’intensité des flux d’azote circulant dans le système. En effet, il semble, d’après les études de cas de ces travaux, que l’intégration agriculture-élevage, bien qu’elle ne se substitue pas aux intrants (autosuffisance), améliore tout de même la productivité globale.
Contrairement aux hypothèses formulées, d’autres relations se dégagent de ces travaux :
- L’autonomie du système semble liée à l’organisation des flux et à la résilience. Nous faisions en effet l’hypothèse que l’autonomie serait liée au niveau d’intégration mais cette relation ne semble pas si évidente à l’échelle d’un système mixte réel. Il semble plutôt que cette autonomie soit liée à la diversité des connexions entre les composantes du système, et ainsi à la résilience globale du système.
- Il en est de même de la relation entre efficience et intensité des flux d’intégration, n’observant pas de substitution entre flux entrants et flux d’intégration, les flux entrants étant l’une des composantes de l’efficience.
Enfin, certaines relations n’ont pu être mises en avant dans ces travaux :
- Les systèmes les plus productifs n’étant pas spécialement les plus consommateurs d’intrants (et vice-versa), la relation entre l’efficience et l’intensité de l’intégration n’apparait pas si évidente.
- La relation entre efficience et résilience n’est pas apparue de façon suffisamment clair dans ces travaux.

Figure 2 : Représentation simplifiée de la projection des études de cas sur le premier plan de l’analyse en composante principale (ACP) fonction des caractéristiques du réseau de flux et des propriétés agroécologiques des systèmes étudiés. Source : Stark, 2016
Analyse de l’effet de la structure paysagère sur les propriétés biologiques (contrôle biologique) et les performances (rendement) de cultures de maïs en Éthiopie
Objectif et problématique
Les systèmes de culture, les pratiques culturales, et la disponibilité d'habitats non cultivés au sein d’un agroécosystème influencent la dynamique spatiale et temporelle des arthropodes dans les paysages agricoles (Bianchi et al., 2013 ; Chateil & Porcher, 2015). En Afrique, le maïs (Zea mays) est une culture vivrière majeure mais sa productivité fluctue considérablement, en relation notamment avec le niveau d’infestation par la pyrale du maïs (Busseola Fusca). Les pratiques de contrôle recommandées se concentrent uniquement à l’échelle de la parcelle et ne prennent pas en compte l'ensemble des caractéristiques du système de production ni la composition du paysage environnant. Cependant, bien que les décisions de gestion agricole soient principalement prises au niveau du champ et de la ferme, la dynamique de la pyrale du maïs et de ses ennemis naturels peuvent être mieux expliqués au niveau du paysage en raison de leur forte mobilité spatiale (Ndjomatchoua et al., 2016). De plus, le contexte paysager peut influencer les interactions ravageurs-ennemis naturels en fournissant des ressources alimentaires et des abris qui varient dans l’espace et le temps (Schellhorn et al., 2014).
Dans cette étude de cas, notre objectif était d’analyser i) comment la structure du paysage et les pratiques agricoles au niveau de la parcelle influencent l'infestation du maïs par la pyrale ainsi que la diversité et l’abondance de ses ennemis naturels, et ii) comment cela impacte in fine les performances du système en termes de rendements du maïs (grain et biomasse) et de contrôle biologique. Les hypothèses majeures sont : (i) la concentration de plantes hôtes de la pyrale du maïs dans un paysage et la réduction des habitats pour leurs ennemis naturels peuvent entraîner une augmentation des taux d'infestation (Root, 1973) ; (ii) plus les taux d’infestation du maïs par la pyrale augmentent, plus les rendements diminuent.
Dispositif de recherche
Dans un premier temps, nous cherchions à comparer des modes de gestion des parcelles de maïs par les producteurs en termes de niveau d’infestation et de rendements. Pour ce faire, nous avons sélectionnés 33 fermes réparties sur trois communes situées dans un gradient de complexité du paysage (Figure 3). Ces paysages présentent une variation de la composition de l’utilisation des sols (proportions variées de cultures annuelles, pérennes et d’habitat non cultivés). Afin de pouvoir comparer les fermes entre elles sans biais lié au contexte paysager, nous avons considéré le pourcentage de la plante hôte principal de la pyrale, le maïs, autour de chaque parcelle à différents radii (100 à 2000 mètres). La caractérisation des pratiques agricoles dans les fermes a été basée sur le suivi de gestion des cultures de maïs durant trois ans (succession de culture, date de semis et variété de maïs, fertilisation azotée du maïs) et les caractéristiques des parcelles (altitude, ratio aire-périmètre, diversité botanique des haies entourant la parcelle, matière organique et réserve phosphorique du sol). Durant trois ans, nous avons relevé les niveaux d’infestation du maïs par la pyrale (en utilisant comme proxy la distance parcourue par la pyrale dans la tige du maïs), et les rendements en grain et biomasse du maïs (Kebede et al., 2019).
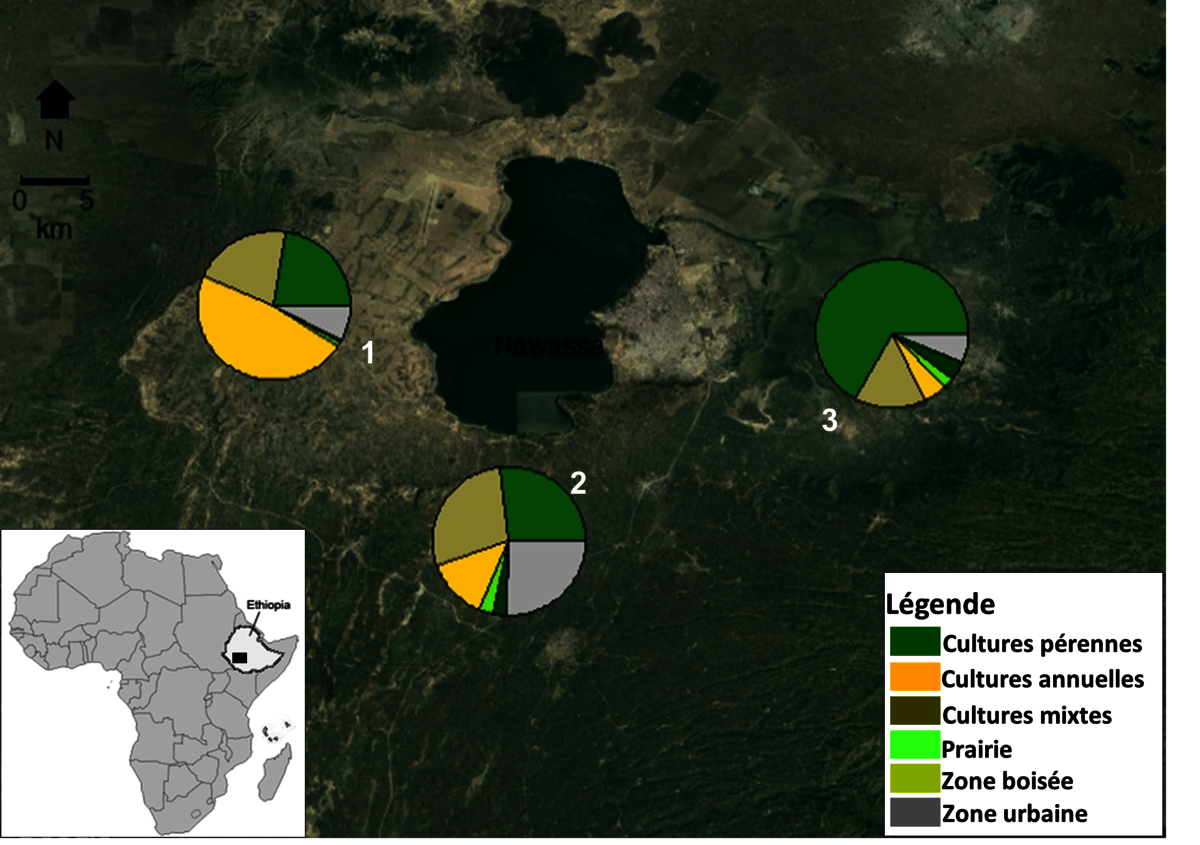
Figure 3 : Localisation des paysages d'étude autour du lac Hawassa dans la vallée du Rift en Éthiopie: paysages simples (1), intermédiaires (2) et complexes (3). Le paysage simple (1) est dominé par le maïs, le paysage diversifié (3) par les cultures pérennes et de la végétation non agricole, et (2) le paysage intermédiaire a une composition mixte de maïs, de cultures pérennes et de végétation non agricole. Source : Kebede, 2019
Dans un second temps, nous avons analysé les performances du système « push-pull », appelé aussi système de répulsion-attraction, une stratégie de culture agroécologique consistant à intercaler la culture d’intérêt avec des plantes attractives d’ennemis naturels et entourées de plantes répulsives d’un ravageur donné. Dans le cas de la lutte contre B. fusca, le maïs est intercalé avec du Desmodium et entouré de Pennisetum purpureum, tous deux étant des cultures fourragères. C’est une stratégie de diversification des cultures prometteuse car elle peut contribuer à contrôler le niveau d’infestation du maïs par B. fusca tout en améliorant la fertilité des sols et en fournissant de l’aliment pour le bétail. Bien que cette stratégie présente des avantages avérés, sa performance n’a été analysée qu’au niveau de la parcelle, sans prendre en compte l’influence du paysage sur la mobilité de la pyrale et de ses ennemis naturels. Dans chacun des trois paysages (de complexité simple, intermédiaire ou complexe), des parcelles expérimentales ont été établies sur quatre exploitations agricoles ; sur chaque exploitation, un dispositif expérimental factoriel en deux blocs et trois parcelles par bloc a été mis en place. Les blocs étaient entourés ou non de P. purpureum et les parcelles secondaires contenaient trois traitements : monoculture de maïs, une culture intercalaire maïs-haricot et une culture intercalaire maïs-Desmodium. Durant deux ans, nous avons analysé les performances de ces différentes variantes du push-pull, en termes de réduction des taux d’infestation, du potentiel d’attraction des ennemis naturels de la pyrale et des rendements du maïs, du haricot, de P. purpureum et du Desmodium (Kebede et al., 2018).
Résultats marquants
La mise en comparaison de différents contextes paysagers et pratiques culturales dans les deux études a permis de tirer trois conclusions majeures :
- les facteurs expliquant l'incidence de la pyrale du maïs varient selon l’échelle considérée (parcelle, ferme et paysage) ;
- la proportion de maïs dans le paysage est le facteur majeur déterminant les taux d’infestation du maïs par la pyrale quelle que soit la pratique au niveau de la parcelle ;
- le rendement de maïs n’est que marginalement affecté par l’infestation quand les taux d'infestation par la pyrale sont faibles à moyens, quel que soit le contexte paysager.
La prise en compte du contexte paysager à différentes échelles autour des 33 fermes a permis de mettre en évidence que le pourcentage de maïs dans le paysage influence le taux d’infestation à des échelles allant de 100 à 1500 m de rayon. Dans les années de forte infestation au niveau régional, les facteurs au niveau de la parcelle et de la ferme n’influencent que marginalement les niveaux d’infestation, même si on note un effet de la densité de semis (l’infestation augmente avec la densité), de la diversité botanique des haies (l’infestation diminue quand la diversité botanique augmente) et de la date de semis (plus le semis est tard, plus l’infestation est élevée).
Le système push-pull s’est révélé efficace pour réduire l'infestation par la pyrale uniquement dans le paysage de complexité intermédiaire, indépendamment du traitement cultural (maïs-P. purpureum, maïs-P. purpureum-haricot ou maïs-P. purpureum-Desmodium) ; les niveaux d’infestation étaient élevés dans les paysages simples dominés par le maïs, et faibles dans les paysages les plus complexes (Figure 4).

Figure 4 : Taux d’infestation de la pyrale de maïs (proxy : longueur des tunnels creusés par la pyrale dans la tige du maïs) par système de production (monoculture maïs (M), maïs-haricot (MB), maïs-Desmodium (MD)) et par gradient de complexité du paysage (simple, intermédiaire, complexe). Source : Kebede, 2019
Principales conclusions
Dans cette étude nous avons orienté les dispositifs expérimentaux vers des approches comparatives afin de pallier la limitation des recommandations de la lutte contre la pyrale du maïs fondées seulement sur des pratiques au niveau de la parcelle, sans prise en compte des effets des paysages environnants. Nous avons montré que la proportion de maïs dans le paysage est le facteur majeur déterminant les taux d’infestation du maïs par la pyrale quelle que soit la pratique au niveau de la parcelle. Ce résultat majeur s’explique par l’hypothèse de concentration des ressources, qui suggère que les insectes herbivores sont plus abondants dans de grandes parcelles de plantes hôtes car ces patchs sont plus faciles à localiser et les herbivores restent plus longtemps dans ces patchs (Root, 1973).
De manière contre-intuitive, les différences de taux d’infestation observées ne se sont pas traduites en une réduction significative des rendements du maïs dans les deux études. Au-delà de l'infestation, la fertilité du sol était le principal facteur de réduction du rendement. Pour le système push-pull, ses avantages proviennent principalement des cultures associées (haricot, Desmodium et P. purpureum) plutôt que de la suppression de la pyrale. De plus, aucune différence significative n’a été trouvée entre les deux cultures intercalaires testées (Desmodium ou haricot) - ce qui suggère que le haricot peut être utilisé comme « push » dans les systèmes push-pull avec l’avantage supplémentaire d’être une pratique courante des producteurs et un apport protéique important.
Analyse de la structure de systèmes de culture bananiers sur leurs propriétés agroécologiques (contrôle biologique) au Costa Rica et en Martinique
Objectif et problématique
Avec l’intensification de l’agriculture, la simplification biologique des agroécosystèmes a entraîné une détérioration de leur fonctionnement écologique, qui a souvent conduit, en substitution, à l’usage d’intrants de synthèse notamment pour lutter contre les ravageurs des cultures (Hooper et al. 2012 ; Vitousek et al. 1997 ; Tilman et al., 2001). De manière croissante, la « re-diversification » des agroécosystèmes est plébiscitée pour restaurer les interactions positives entre les plantes au bénéfice de la culture et réduire le recours à la pétrochimie (Birch et al. 2011 ; Lewis et al. 1997 ; Zhao et al. 2016). Mais quelle devrait être la configuration de la communauté végétale à l’échelle de la parcelle pour favoriser la résistance des agroécosystèmes aux ravageurs ? Telle est la question à laquelle ce travail (Poeydebat, 2017) tentait de répondre.
Plus particulièrement, notre objectif était de déterminer et quantifier le rôle de la configuration de la communauté végétale à l’échelle de la parcelle dans la résistance des bananiers à trois types de ravageurs, le charançon, la maladie des raies noires et la cohorte de nématodes endoparasites du bananier (Figure 5). Il était également question de déterminer quelles composantes de la végétation jouaient un rôle dans les différents cas et d’inférer les mécanismes écologiques à l’œuvre. In fine, l’objectif était d’identifier quelle devraient être la composition et l’arrangement spatial de la communauté végétale dans la parcelle pour optimiser de façon synergique la résistance des agroécosystèmes bananiers à de multiples ravageurs.
Dispositif de recherche
Dans le cas présent, l’approche comparative a consisté à mettre en regard des agrosystèmes bananiers variés en termes de diversité végétale et à comparer leurs propriétés de résistance à différents ravageurs.
Pour cela, nous avons sélectionné 19 parcelles de producteurs de bananes en Martinique et au Costa Rica, de sorte à constituer un large gradient de diversité végétale dans un pédo-climat relativement homogène. Dans chaque parcelle, nous avons délimité une ou plusieurs placettes circulaires de dix mètres de rayon, et au centre de chacune d’elles nous avons implanté un bananier de type Cavendish faisant office de phytomètre. Nous avons suivi un total de 124 placettes (ou phytomètres) dont la richesse végétale variait de 1 à 32 espèces (Figure 5).
Dans chacune des placettes, nous avons caractérisé la structure de l’agrosystème bananier en mesurant différentes composantes de la végétation (notamment richesse spécifique de différentes strates, richesse génotypique des Musa, type de couverture du sol, ouverture de la canopée), ainsi que l’abondance de différents groupes d’organismes du sol et du sous-sol (arthropodes, nématodes) et le pédo-climat. Enfin, nous avons évalué l’abondance des trois ravageurs dans les placettes ainsi que les dégâts sur les phytomètres.
En utilisant des méthodes adaptées dans chaque cas (modèles linéaires, courbes enveloppes ou différence entre niveaux potentiels et observés), nous avons tout d’abord estimé l’effet de la richesse spécifique de la communauté végétale dans la parcelle sur l’abondance et les dégâts des trois ravageurs. Grâce à la méthode des équations structurelles (Grace, 2006), nous avons ensuite pu tester l’effet de différentes composantes de la structure de la communauté végétale dans la parcelle sur l’abondance et les dégâts des trois ravageurs, mais aussi déterminer si ces effets étaient directs ou indirects (via des cascades trophiques ou des interactions biotiques indirectes). Notre objectif était de tester différentes hypothèses concernant les mécanismes sous-jacents.
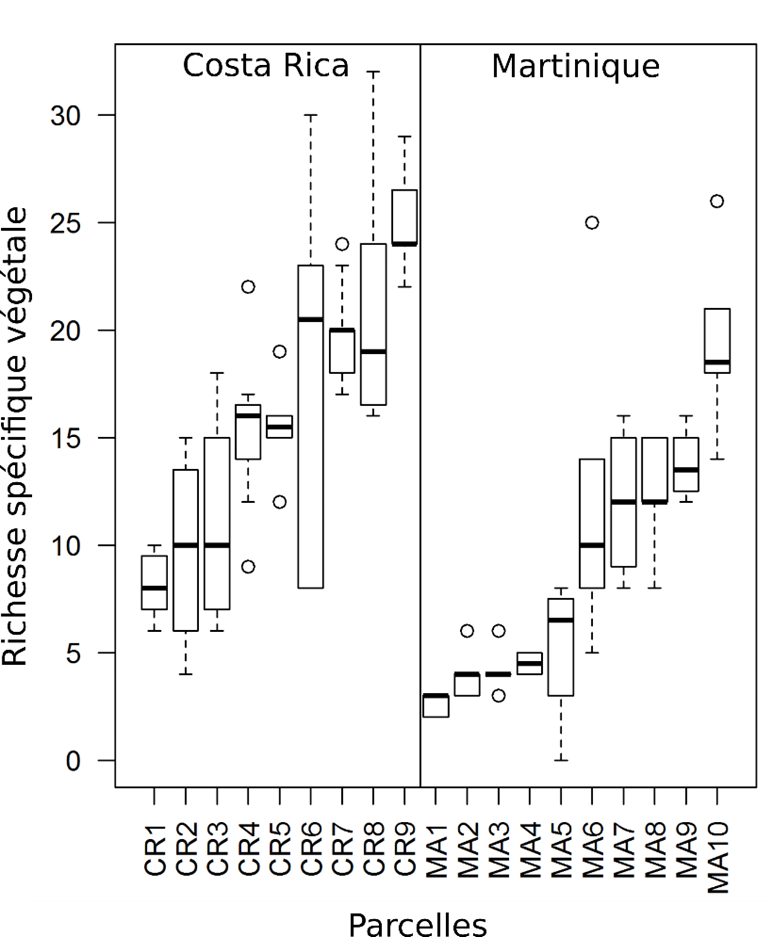
Figure 5 : Graphiques en boîte à moustache montrant la richesse spécifique végétale dans les placettes sélectionnées pour l’étude, d’une part (à gauche) au Costa Rica et d’autre part (à droite) en Martinique. On constate que les placettes constituent un gradient de diversité végétale important. Cette variabilité du niveau de diversité était nécessaire à l’étude. Source : Poeydebat, 2016
Résultats marquants
Premièrement, nous avons montré que la richesse spécifique de la communauté végétale dans la parcelle jouait un rôle dans le contrôle de la maladie des raies noires et du charançon du bananier.
En cherchant à décortiquer l’effet de la structure de la communauté végétale, nous avons montré que l’abondance des nématodes parasites du bananier augmentait avec la richesse variétale des bananiers dans la parcelle (plus il y a de variétés, plus il y a de chances d’en inclure des particulièrement susceptibles) et que l’abondance des charançons augmentait avec l’abondance de bananiers dans la parcelle (hypothèse de concentration de la ressource).
La richesse de la strate herbacée ainsi que la proportion de sol couvert de débris végétaux favorisaient l’abondance et la diversité de proies alternatives et d’ennemis naturels du charançon et des nématodes, ennemis qui étaient impliqués dans la régulation des deux ravageurs par prédation ou antagonisme.
L’abondance de spores de Mycosphaerella fijiensis (l’agent de la maladie des raies noires) atteignant le feuillage des bananiers (effet barrière) et l’efficacité d’infection de la maladie des raies noires (temps d’incubation plus long et densité de lésions réduite ; microclimat et microorganismes épiphylles antagonistes) avaient tendance à diminuer avec l’augmentation de la richesse végétale entre 1,5 et 5 mètres de hauteur.
De ces résultats (Figure 6), nous avons tenté de dégager une vision globale et d’identifier les synergies et antagonismes dans les effets des composantes de la structure de la communauté végétale. Sur cette base, nous avons fait une série de propositions concernant la gestion de la communauté végétale à l’échelle de la parcelle pour favoriser la résistance des bananeraies à de multiples ravageurs :
- mettre en place une matrice de type de couverts du sol (sol nu, débris végétaux et couvert vivant) pour favoriser l’abondance de proies alternatives herbivores et détritivores ainsi que l’activité des ennemis naturels ;
- au sein des patches de couvert vivant, promouvoir la richesse spécifique (pas seulement la biomasse) pour supporter une plus grande abondance et diversité de proies alternatives, tout en excluant les espèces hôtes des ravageurs généralistes (les nématodes) ;
- diversifier la végétation dans la strate haute (hauteur des bananiers) pour modifier le microclimat, favoriser la diversité des cohortes de microorganismes épiphylles, et enfin perturber l’accès des ravageurs aériens à leurs hôtes (effet barrière) ;
- limiter la densité de bananiers ;
- choisir des cultivars résistants.

Figure 6 : Schéma synthétisant les résultats explicitant les liens entre diversité végétale et abondance et dégâts des trois ravageurs. Source : Poeydebat, 2016
Principales conclusions
Ces travaux ont permis d’estimer les services rendus par la diversité végétale en termes de régulation des ravageurs des bananiers et aussi d’inférer les mécanismes écologiques sous-jacents. La somme des résultats mis en évidence pour chacun des ravageurs montre que le système étudié est relativement complexe. Cependant nous avons montré qu’il était possible de le décomposer pour le comprendre et ensuite d’en reconstruire une image globale. Il ressort que la capacité des systèmes bananiers à résister à de multiples ravageurs est fortement liée à la structure de la communauté végétale à l’échelle de la parcelle.
Discussion générale et conclusion
Par rapport aux initiatives visant à mettre en commun des données sur les performances de différents systèmes agricoles, (par exemple Sachs et al., 2010), sur leur positionnement édaphique (Tóth et al., 2020) et à celles qui traitent de problématiques globales (par exemple liens entre activités agricoles et changement climatique, ou activités agricoles et capacité nourricière, voir Licker et al., 2010) les trois études présentées partagent les caractéristiques suivantes :
- Elles visent à recenser non seulement des performances mais aussi à expliquer les relations entre les composantes d’un système faites de relations directes et indirectes pas toujours appréhendables deux à deux ; entre des caractéristiques et des propriétés des agroécosystèmes pour une diversité de contextes, dans une démarche davantage explicative ;
- Elles misent sur les bénéfices heuristiques que l’on peut attendre de comparaisons approfondies d’études de cas, de manière significativement plus ambitieuse qu’à travers un simple parangonnage, pour décortiquer les mécanismes sous-tendant les relations structure-propriété ;
- Elles complètent les approches globales en se positionnant dans une dynamique prospective du développement agricole, en visant à identifier les propriétés pertinentes à rechercher dans les agroécosystèmes.
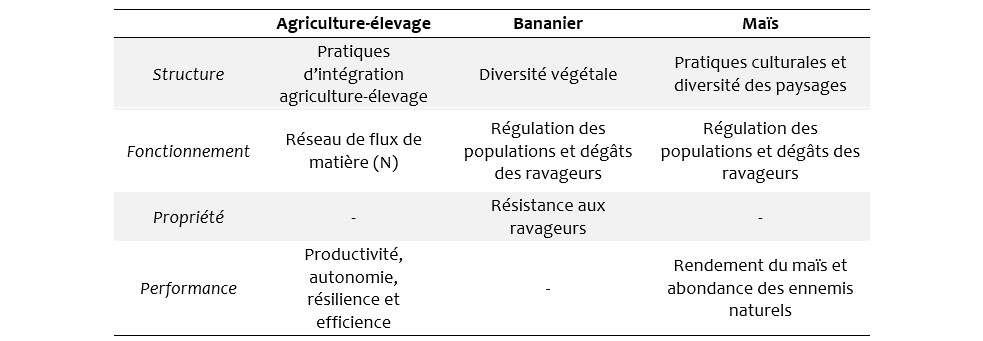
Tableau 2 : Récapitulatif des trois études de cas dans une grille « agronomie comparative »
Même si elles investissent différemment les relations structure – propriétés – performances, ces trois études de cas présentées dans cet article montrent certaines similitudes dans les démarches mise en œuvre (Tableau 2). Bien qu’elles n’aient pas été conçues en utilisant le référentiel structure/propriétés/performances, il s’agit bien de travaux qui permettent d’évaluer les performances attendues de la part d’agroécosystèmes, suivant différentes caractéristiques liées à des modalités de gestion. Plus que l’évaluation des systèmes en tant que telle, ces travaux cherchent à expliciter les relations entre la structure des systèmes étudiés, leur fonctionnement, les propriétés et in fine les performances associées, sur une base comparative. Ainsi, les trois études de cas donnent corps à l’idée d’agronomie comparative, en cousinage avec ce qui se pratique dans d’autres disciplines. L’anatomie comparée, par exemple, consiste à comparer l’anatomie de différentes espèces pour en déterminer la phylogénie et identifier les processus adaptatifs ayant œuvré dans différents environnements. Cela revient à comprendre l’aspect fonctionnel de caractéristiques anatomiques des espèces. Par analogie, la biologie comparative utilise les variations et disparités naturelles pour comprendre les modes du vivant à tous les niveaux (des gênes aux communautés) et le rôle (la fonction) des organismes dans les écosystèmes. Par extension, on peut dire que l’agronomie comparative peut utiliser la variabilité des situations agricoles existantes pour comprendre comment leur configuration (structure) influence leur fonctionnement et donc les propriétés des agroécosystèmes, avec des impacts sur leurs performances. Bonaudo et al. (2016) proposent par exemple, en s’inspirant du cadre conceptuel de l’agroécologie, de mettre en relation ce qu’ils appellent propriétés émergentes (résilience, efficience, autosuffisance et productivité), principes écologiques (diversité et interactions) et pratiques agricoles (fonctionnement).
La volonté explicative de l’approche est bien présente dans les trois travaux considérés : dans le cas de l’étude sur les systèmes bananiers par exemple, il ne s’agit pas uniquement de mesurer la capacité des bananiers à résister à des bioagresseurs en fonction du niveau de diversité végétale dans la parcelle, mais de comprendre par quels mécanismes écologiques la diversité végétale confère ou pas cette résistance aux bananiers. Ainsi, différentes hypothèses sur les effets de la diversité végétale sont testées grâce à la prise en compte d’éléments intermédiaires (réseaux trophiques, micro-climat, propriétés du sol). Il s’agit aussi de travaux fondamentalement systémiques à dimension heuristique, pour lesquels l’ensemble des hypothèses n’est pas formulé explicitement mais découle de cette mise en comparaison. Dans le cas des travaux sur les systèmes agriculture-élevage par exemple, des approches issues de l’écologie sont mobilisées (analyse de réseau écologique) pour identifier les propriétés émergentes de ces systèmes, fruits des relations indirectes entre les composantes du système. Il est dès lors possible de mettre en relation la résilience du système étudié et la « redondance » des flux de matière au sein du système, par la mise en comparaison de situations différentes, difficilement appréhendable de manière segmentée. Enfin, dans une vision dynamique des activités de recherche et de développement, ces travaux permettent de faire émerger des propriétés pertinentes qu’il reste ensuite nécessaire d’approfondir pour monter en généricité. Il s’agit en effet de dispositifs qui présentent une certaine richesse du fait de la gamme de situations analysée suivant une approche commune, mais de situations qui présentent aussi une forte variabilité sur tout un ensemble de facteurs. Dès lors, il peut s’avérer pertinent de prolonger le dispositif comparatif par des travaux analytiques plus ciblés pour préciser les propriétés identifiées et les conditions associées. Les travaux de l’étude sur maïs montrent par exemple l’influence de la complexité paysagère sur le niveau d’infestation de la pyrale du maïs, quelles que soient les pratiques mises en œuvre à l’échelle de la parcelle. Ces résultats questionnent ainsi l’échelle à laquelle gérer certaines maladies et la complémentarité entre ces échelles pour des modes de gestion durables et efficaces.
Les trois études de cas de cet article se sont toutes appuyées sur des gradients de situations différentes pour pouvoir comparer des processus s’exprimant de manière différenciée, en fonction de ces situations. Dans ce type de travaux à dimension comparative, l’échantillonnage correspond en effet à un gradient de situations qui se construit différemment suivant la problématique posée et à différentes échelles. Ces études permettent d’interroger ce que serait une stratégie d’échantillonnage en agronomie comparative. La première question concerne la nature du gradient : que compare-t-on ? Sur quelle base ? La complexité des processus étudiés, s’inscrivant dans des situations réelles aux interactions multiples, nécessite en effet d’avoir recours à des proxys pour constituer un échantillon à même de répondre à la question posée. Dans le cas des travaux sur bananier, il s’agit d’un gradient de diversité végétale au niveau de la parcelle, dans le cas des travaux sur agriculture-élevage, le gradient est construit en fonction d’une gamme de structures d’exploitations en polyculture-élevage, et dans le cas des travaux sur maïs, suivant un gradient de complexité paysagère. Pour autant, les échelles constitutives du gradient de situations ne correspondent pas forcément au processus étudié ; les objets d’études de ces travaux se situant à des échelles infras : ravageurs pour le bananier et le maïs, pratiques agricoles pour agriculture-élevage. Il s’agit en effet d’approches systémiques pour lesquelles le processus étudié est le fruit de processus biologiques sous-jacents et s’inscrivant dans des interactions à des échelles supérieures. Dans le cas des travaux sur bananier par exemple, il s’agit d’étudier le niveau d’abondance et de dégât de ravageurs, fonction des conditions biologiques plus ou moins propices à leur développement, favorisé par la structure de la communauté végétale (diversité végétale parcellaire).
Une autre question relative à ce gradient de situations concerne la variabilité attendue pour rendre compte du processus étudié et de ses implications. Il est en effet important d’avoir recours à des situations suffisamment contrastées pour rendre compte de processus s’exprimant différemment suivant la structure des objets étudiés, et conduisant à des performances qui peuvent être différentes. Le choix peut alors se porter vers des situations « extrêmes », très contrastées, et/ou vers des situations « graduelles » permettant de rendre compte de manière plus fine des processus à l’œuvre. Dans le cas des travaux sur agriculture-élevage, par exemple, la constitution d’un gradient initial fondé uniquement sur des situations Guadeloupéennes n’avait pas permis de montrer de différences notables en matière de réseau de flux de nutriments et de performances associées. Le recours à un échantillonnage plus large (Guadeloupe-Amazonie brésilienne-Cuba) a ainsi permis d’élargir cette gamme et de mettre en exergue des relations entre réseau de flux et performances de ces systèmes polyculture-élevage. Il en va de même pour les travaux sur bananier, où les deux zones d’échantillonnage (Martinique et Costa Rica) ont été choisies pour élargir le gradient de diversité végétale dans les parcelles, les parcelles costariciennes offrant un plus haut niveau de diversité
Au même titre que le chercheur en anatomie comparée caractérise la morphologie des espèces qu’il étudie pour pouvoir la comparer, l’agronome qui cherche à mettre en œuvre une analyse comparative d’agroécosystèmes devra être en mesure d’identifier les indicateurs pertinents ou d’intérêt pour caractériser ces systèmes et ce qui, en les différenciant, rend possible la comparaison. A contrario d’approches expérimentales en situation contrôlée, le principe « toutes choses étant égales par ailleurs » est impossible à suivre dans ce type de dispositifs comparatifs. Pour autant, le gradient ainsi constitué doit permettre de comparer des processus comparables, suivant des facteurs de variations préalablement identifiés. Telle est l’une des fonctions des proxys, fondés sur les hypothèses formulées. L’étude sur maïs fait en effet l’hypothèse que le taux d’infestation du maïs par la pyrale est fonction de la proportion de plante hôte à une échelle plus large que la parcelle, d’où un gradient de complexité paysagère plutôt qu’un gradient de maïs à l’échelle parcellaire uniquement.
Finalement, les approches comparatives en agronomie sont à même de contribuer aux réponses apportées par la recherche pour répondre aux enjeux agricoles à différents niveaux. Vis-à-vis des changements à l’œuvre tel que le changement climatique, les approches comparatives suivant des gradients de situations environnementales pourraient permettre d’identifier des pistes d’adaptations et de comprendre l’effet de ces changements sur les processus agronomiques. Vis-à-vis des enjeux de diversification des systèmes agricoles pour fournir des services écosystémiques, comparer des systèmes aux niveaux de diversité variés et comprendre l’effet de la diversité sur le fonctionnement de l’agroécosystème pourraient permettre de formuler des connaissances génériques sur l’effet de la diversification. L’agronomie, dans le cadre de l’agroécologie notamment, tend à s’appuyer de plus en plus sur les cadres conceptuels de l’écologie pour chercher à comprendre les interactions fines entre les composantes biotiques et abiotiques des systèmes cultivés, par ailleurs appelés à être plus diversifiés biologiquement à différentes échelles et au fonctionnement écologique complexe. La prise en considération de situations extrêmes et pas uniquement des situations moyennes, à travers des gradients de situation est aussi une entrée intéressante des approches comparatives pour identifier des situations et des processus pouvant présenter un intérêt pour concevoir de nouvelles formes de systèmes agricoles. Cependant, les cas d’étude présentés ici montrent que l’approche comparative des agroécosystèmes pose plusieurs questions méthodologiques. À quelle échelle comparer les systèmes ? Jusqu’à quel point doit-on y intégrer des composantes humaines, et traiter le cas échéant de socio-agroécosystèmes ? Quels éléments structurels est-il intéressant de caractériser ? Quelles propriétés sont à privilégier ? Quelles relations entre propriétés (de fonctionnement) et processus ? Une démarche formalisée d’agronomie comparative devrait s’attacher à répondre à ces questions et à en préciser les théories sous-jacentes.
Références bibliographiques
Berger, J., Robertson, L., Cocks, P., 2002. Agricultural potential of Mediterranean grain and forage legumes: Key differences between and within Vicia species in terms of phenology, yield, and agronomy give insight into plant adaptation to semi-arid environments. Genetic Resources and Crop Evolution, 49, 313–325. doi.org/10.1023/A:1015544126185
Bianchi, F.J.J.A., Ives, A., Schellhorn, N., 2013. Interactions between conventional and organic farming for biocontrol services across the landscape. Ecol Appl 23, 1531-1543
Birch A.N.E., Begg G.S., Squire G.R., 2011. How agro-ecological research helps to address food security issues under new IPM and pesticide reduction policies for global crop production systems. Journal of Experimental Botany 62:3251-3261.
Bonaudo T., Bendahan A.B., Sabatier R., Ryschawy J., Bellon S., Leger F., Magda D., Tichit M., 2014. Agroecological principles for the redesign of integrated crop–livestock systems. Eur J Agron 57, 43-51.
Cann D.J., Schillinger W.F., Hunt J.R., Porker K.D., Harris F.A.J., 2020. Agroecological Advantages of Early-Sown Winter Wheat in Semi-Arid environments: A Comparative Case Study From Southern Australia and Pacific Northwest United States. Front. Plant Sci. 11:568. doi: 10.3389/fpls.2020.00568
Chateil C., Porcher E., 2015. Landscape features are a better correlate of wild plant pollination than agricultural practices in an intensive cropping system. Agriculture, Ecosystems & Environment 201, 51-57.10.1016/j.agee.2014.12.008
Doré T., 2010. L’agronomie demain. Cahiers agricultures, 19(3), 175-176. DOI : 10.1684/agr.2010.0407
Doré T., Makowski D., Malézieux E., Munier-Jolain N.G., Tchamitchian M., Tittonell P., 2011. Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. European Journal of Agronomy, 34 (4), pp.197-210. DOI : 10.1016/j.eja.2011.02.006
Fuhrman J.A., Steele J.A., 2008. Community structure of marine bacterioplankton: patterns, networks, and relationships to function. Aquat. Microb. Ecol., 53, 69-81.
Garnier E. et al., 2007. Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: A standardized methodology and lessons from an application to 11 European sites. Annals of botany, 99(5), 967-985. doi:10.1093/aob/mcl215
González-García E., Gourdine J.L., Alexandre G., Archimede H., Vaarst M., 2012. The complex nature of mixed farming systems requires multidimensional actions supported by integrative research and development efforts. Animal 6, 763–777.
Grace J.B., 2006. Structural equation modeling and natural systems. Cambridge University Press.
Holzkämper A., Calanca P., Fuhrer J., 2011. Analyzing climate effects on agriculture in time and space. Procedia Environmental Sciences, 3, pp.58-62.
Hooper D.U. et al., 2012. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486:105-108.
Huey R.B., Garland T., Turelli M., 2019. Revisiting a Key Innovation in Evolutionary Biology: Felsenstein’s “Phylogenies and the Comparative Method”. The American Naturalist, 2019, 193:6, 755-772. DOI: 10.1086/703055
Kebede Y., Baudron F., Bianchi F.J.J.A., Tittonell P., 2018. Unpacking the push-pull system: Assessing the contribution of companion crops along a gradient of landscape complexity. Agriculture, Ecosystems & Environment, 268, pp.115-123. doi.org/10.1016/j.agee.2018.09.012.
Kebede Y., 2019. Hide and seek: management and landscape factors affecting maize stemborers Busseola fusca (Fuller) infestation levels in Ethiopia (Doctoral dissertation, Wageningen University).
Kebede Y., Bianchi F.J.J.A., Baudron F., Tittonell P., 2019. Landscape composition overrides field level management effects on maize stemborer control in Ethiopia. Agriculture, Ecosystems & Environment, 279, pp. 65-73.
Lewis W.J., Van Lenteren J.C., Phatak S.C., Tumlinson Lii J.H. 1997. A total system approach to sustainable pest management. Proceedings of the National Academy of Sciences of the United States of America 94:12243-12248.
Licker R., Johnston M., Foley J.A., Barford C., Kucharik C.J., Monfreda C. and Ramankutty, N., 2010., Mind the gap: how do climate and agricultural management explain the ‘yield gap’ of croplands around the world? Global Ecology and Biogeography, 19, 769-782. doi:10.1111/j.1466-8238.2010.00563.x
Mauseth J.D., 2006. Structure–function relationships in highly modified shoots of Cactaceae. Ann. Bot., 98, 901-926.
Ndjomatchoua F.T., Tonnang H.E.Z., Plantamp C., Campagne P., Tchawoua C., Le Ru B.P., 2016. Spatial and temporal spread of maize stemborer Busseola fusca (Fuller) (Lepidoptera: Noctuidae) damage in smallholder farms. Agriculture, Ecosystems & Environment 235, 105-118.http://dx.doi.org/10.1016/j.agee.2016.10.013
Poeydebat C., 2016. Régulations écologiques d'un complexe de bioagresseurs du bananier dans un gradient de complexité végétale des agroécosystèmes. Thèse de doctorat, Université de Montpellier.
Root R.B., 1973. Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica Oleracea). Ecological Monographs 43, 95-124
Rosencrans R.F., Leslie C.E., Perkins K.A., Walkowski W., Gordon W.W., Richards-Zawacki C.L., Bazan N.G., Farris H.E., 2018.Quantifying the relationship between optical anatomy and retinal physiological sensitivity: a comparative approach. J. of comparative neurology, 526(18), 3045-3057. doi: 10.1002/cne.24531
Rufino M.C., Hengsdijk H., Verhagen A., 2009. Analysing integration and diversity in agro-ecosystems by using indicators of network analysis. Nutr. Cycl. Agroecosyst. 84, 229–247.
Sachs J. et al., 2010. Monitoring the world's agriculture. Nature, 466(7306), 558-560. doi:10.1038/466558a
Schellhorn N.A., Bianchi F.J.J.A., Hsu C.L., 2014. Movement of entomophagous arthropods in agricultural landscapes: links to pest suppression. Annual review of entomology 59, 559- 581.10.1146/annurev-ento-011613-161952
Schluessel V., Bennett M.B., Bleckmann H., Blomberg S., Collin, S.P. (2008), Morphometric and ultrastructural comparison of the olfactory system in elasmobranchs: The significance of structure–function relationships based on phylogeny and ecology. J. Morphol., 269: 1365-1386. doi:10.1002/jmor.10661
Stark F., 2016. Évaluation agroécologique des pratiques d’intégration agriculture-élevage: Application de l’analyse de réseau écologique à des exploitations agricoles en milieu tropical humide. Thèse de doctorat, AgroParisTech, 246 p.
Stark F., Archimède H., González-García E., Poccard–Chapuis R., Fanchone A., Moulin C.H., 2019. Evaluation des performances agroécologiques des systèmes de polyculture- élevage en milieu tropical humide : application de l’analyse de réseaux écologiques, Innovations Agronomiques, 72, 1-14
Thomson K.S., Simanek D.E., 1977. Body Form and Locomotion in Sharks, American Zoologist, 17(2) 343–354, https://doi.org/10.1093/icb/17.2.343
Tilman D., et al., 2001. Forecasting agriculturally driven global environmental change. Science 292:281-284.
Tóth G., Kismányoky T., Kassai P., Hermann T., Fernandez-Ugalde O., Szabó B., 2020. Farming by soil in Europe:status and outlook of cropping systems under different pedoclimatic conditions. PeerJ 8:e8984 http://doi.org/10.7717/peerj.8984
Ulanowicz R.E., Goerner S.J., Lietaer B., Gomez R., 2009. Quantifying sustainability: Resilience, efficiency and the return of information theory. Ecol. Complex. 6, 27–36.
Vitousek P.M., Mooney H.A., Lubchenco J., Melillo J.M. 1997. Human domination of Earth's ecosystems. Science 277:494-499.
Zhao Z.H., Reddy G.V.P., Hui C., Li B.L. 2016. Approaches and mechanisms for ecologically based pest management across multiple scales. Agriculture, Ecosystems and Environment 230:199-209.
Zink R.M., 2002. Methods in Comparative Phylogeography, and Their Application to Studying Evolution in the North American Aridlands, Integrative and Comparative Biology, 42(5) 953–959

Les articles sont publiés sous la licence Creative Commons (CC BY-NC-ND 2.0)
Pour la citation et la reproduction de cet article, mentionner obligatoirement le titre de l'article, le nom de tous les auteurs, la mention de sa publication dans la revue AES et de son URL, la date de publication.