

L’agriculture régénératrice, une démarche pour massifier la transition vers l’agroécologie
Olivier Husson*, Jean-Pierre Sarthou**, Olivier Thérond*** et Michel Duru
* CIRAD, UPR AIDA, Avenue Agropolis F34398, Montpellier
** AgroToulouse INP, UMR CRBE, Université de Toulouse
*** INRAE, UMR 1132 LAE, 28 rue de Herrlisheim, 68 000 Colmar
contact auteurs : michel.duru@gmail.com
Résumé
L’agroécologie repose en premier lieu sur la promotion de la biodiversité dans les agroécosystèmes pour faire face à de multiples défis. Massifier son développement est cependant difficile car passer des principes aux pratiques suppose de tenir compte des spécificités des situations (sol, climat, histoire culturale) et de définir une trajectoire pour combiner de manière cohérente sur le temps long l’accroissement de la biodiversité dans les cultures, les sols et les paysages, et l’utilisation de plus en plus parcimonieuse d’intrants d’origine fossile. Le concept de régénération des écosystèmes cultivés permet de définir cette trajectoire de développement des services écosystémiques et de réduction progressive des intrants, et ainsi de trouver un écho auprès d’une grande diversité d’acteurs. Après avoir rappelé les processus écologiques sous-jacents à la fourniture de services écosystémiques pouvant remplacer des intrants, nous formulons les bases d’une stratégie de régénération des agroécosystèmes dégradés et/ou simplifiés, ou reposant sur une utilisation d’intrants excessive non compatible avec les objectifs des politiques publiques. Pour ces cas de figures, elle repose, après diagnostic précis de l’agroécosystème, sur le remplacement progressif des apports directs (carburants) et indirects (engrais et pesticides de synthèse) en énergie fossile, par l’énergie de la photosynthèse, directement et indirectement, ainsi que sur des interventions humaines favorables à la biodiversité fonctionnelle bénéfique.
Mots clefs : agriculture biologique ; agriculture de conservation des sols ; biodiversité ; potentiel redox ; santé du sol ; services écosystémiques ; transition agroécologique
Abstract
Regenerative agriculture: a pathway to scaling up the agroecological transition?
Agroecology is primarily based on promoting biodiversity in agroecosystems to address multiple challenges. However, development is difficult because moving from principles to practices requires taking into account the specificities of situations (soil, climate, crop history) and defining a trajectory to consistently combine over the long term the increase in biodiversity in crops, soils and landscapes, and the decrease in fossil-based inputs. The concept of regeneration of agroecosystems makes it possible to define this trajectory of providing of ecosystem services and gradual reduction of inputs, and thus to resonate with a wide variety of stakeholders. After recalling the ecological processes underlying the provision of ecosystem services that can replace inputs, we formulate the bases of a strategy for regenerating degraded and/or simplified agroecosystems, or those based on excessive use of inputs that is incompatible with public policy objectives. After precise diagnosis of agroecosystem state, it is based on the progressive replacement of direct (fuels) and indirect (synthetic fertilizers and pesticides) inputs of fossil energy, by the energy of photosynthesis, directly and indirectly, as well as on human interventions favorable to beneficial functional biodiversity.
Key words: agroecological transition; biodiversity; ecosystem services; organic farming; soil conservation agriculture; redox potential; soil health
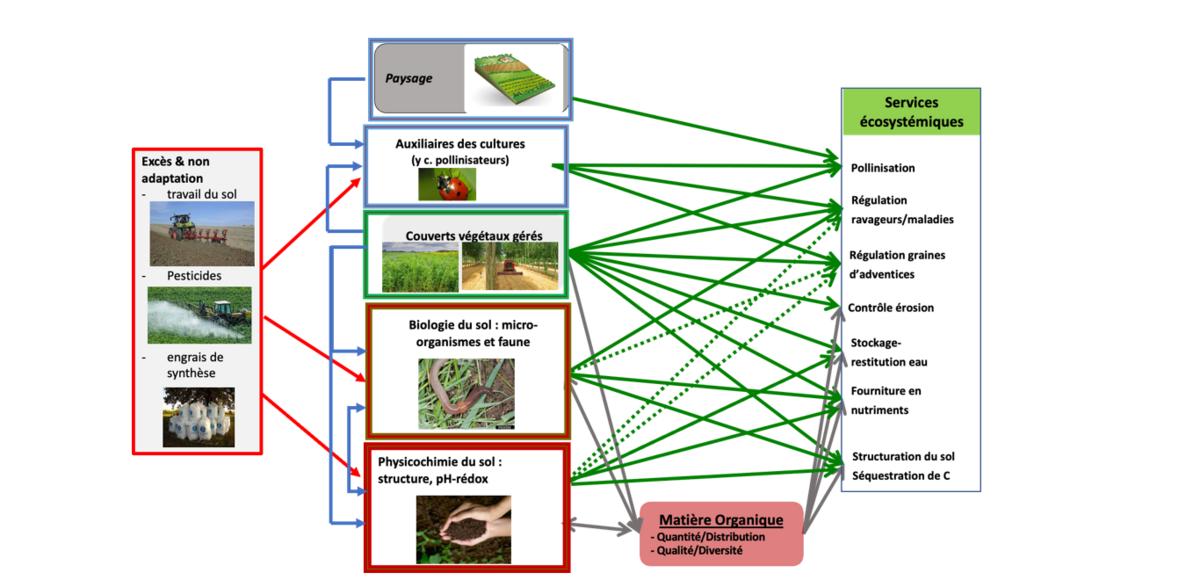
Flèches rouges : effets négatifs des pratiques sur la biodiversité
Flèches bleues : relations positives entre biodiversité intentionnelle et associée
Flèches vertes : effets de la biodiversité sur la fourniture de services écosystémiques
Flèches grises : relations entre biodiversité et services via la MO
En pointillé, flèches dont le lien n’est pas encore pleinement démontré
Transitionner vers une agriculture régénératrice : enjeux et difficultés
Au cours des cinquante années qui ont suivi la Seconde Guerre mondiale, les rendements de l’agriculture européenne ont fortement augmenté du fait d’un accroissement de l’utilisation des intrants de synthèse et de l’amélioration génétique (Rigal et al., 2023 ; Seibold et al., 2019). Il en a été de même de la productivité du travail, grâce à l’énergie fossile surtout, du fait de la mécanisation, de l’agrandissement des parcelles, et de la spécialisation et standardisation des modes de production (Steenwyk et al., 2022).
Depuis les années 2000, il est cependant acquis que cette évolution s’est faite au détriment de l’environnement : érosion, pollutions par les pesticides et les composés azotés, raréfaction des ressources (énergie, eau, phosphore) (Prăvălie et al., 2024), déclin de la biodiversité sauvage et cultivée (agrandissement des parcelles, disparition des haies, spécialisation des productions…) (Newbold et al., 2015). De plus en plus de travaux démontrent aussi l’impact négatif des pesticides sur notre santé (Kesse-Guyot et al., 2017).
Pour faire face à ces impacts environnementaux, deux grandes formes de transition agricole ont émergé (Duru et al., 2022). L’une, agriculture raisonnée, vise la réduction des impacts grâce aux technologies d’optimisation de l’utilisation des intrants de synthèse, sans remettre en cause la simplification des systèmes agricoles. L’autre, appelée agroécologie, est basée sur la gestion de la biodiversité qui sous-tend les services écosystémiques. Elle représente un véritable changement de paradigme puisqu’il s’agit de passer d’un modèle d’optimisation, centré sur l’apport du bon produit au bon endroit, au bon moment et à la bonne dose, mobilisant beaucoup de ressources industrielles, à un modèle mobilisant le vivant de façon agile via l’adaptation des pratiques à l’état de l’agroécosystème, de façon à remplacer autant que possible les intrants de synthèse par les services que fournit la biodiversité fonctionnelle bénéfique (ex. fourniture de nutriments, contrôle biologique des ravageurs, pollinisation…) (Duru et al., 2015).
Les formes d’agriculture se revendiquant de l’agroécologie, telles que l’agriculture biologique (AB), l’agriculture de conservation des sols (ACS), l’agroforesterie (AF) ou encore l’agriculture régénératrice (AR), sont le plus souvent apparues en réaction aux impacts environnementaux de l’agriculture conventionnelle (Duru et al., 2022). Un de leurs points communs est d’accorder de l’importance à la matière organique (MO) du sol et à la biodiversité planifiée (espèces cultivées ou élevées) et associée (organismes du sol, auxiliaires comme les ennemis naturels des ravageurs et les pollinisateurs) ; deux caractéristiques centrales pour augmenter les services écosystémiques (SE) à l’agriculture (réduction des intrants de synthèse et des énergies directes et indirectes) et à la société (multiples bénéfices environnementaux) (Therond et Duru, 2019). Alors que de nombreuses recherches sont conduites sur l’AB (Carné-Carnavalet, 2011) et l’ACS (Cordeau et al., 2024), l’AR est surtout l’apanage de l’agro-industrie et n’a pas de définition stabilisée (Tittonell et al., 2022). En outre, la distinction avec l’agriculture dite conventionnelle (AC) n’est pas toujours facile car de plus en plus d’agriculteurs mettent en place, plus ou moins ponctuellement, spatialement et temporellement, des principes agroécologiques tels que des couverts végétaux en interculture, des mélanges variétaux et associations de cultures, du semis direct ou des méthodes de biocontrôle y compris par la mise en place d’infrastructures agroécologiques. Cependant, ces systèmes agroécologiques présentent plusieurs limites : les rendements et le travail du sol en AB, l’utilisation encore élevée d’herbicides en ACS, le temps d’entretien des jeunes arbres en AF. L’AR, surtout portée par l’agro-industrie, est aussi souvent qualifiée de greenwashing (Tittonell et al., 2022 ; Duru et al., 2022).
Dépasser ces limites dans une perspective de massification des systèmes agroécologiques, nécessite d’une part de prendre en compte la situation de départ : degré de dégradation des sols et de simplification des systèmes agricoles et paysages, utilisation excessive d’intrants de synthèse non compatible avec les objectifs de politiques publiques, et d’autre part de définir des principes et étapes pour transitionner. Cette transition vise à accroître la fourniture de services à la société (séquestration de carbone, préservation de l’eau, de la biodiversité, des paysages…) en améliorant/restaurant certaines fonctionnalités écologiques et, par ricochet, à réduire les impacts (résidus de pesticides, émissions d’azote et de gaz à effet de serre). À cet effet, le concept de régénération est plus qu'un simple mot à la mode. La régénération des écosystèmes cultivés s'impose comme un objet-frontière essentiel d'un changement de paradigme qui redéfinit les principes de conception pour passer du seul objectif de réduction des impacts, à la fourniture de services (Paolini et al., 2024). C’est bien aussi le fondement de l’agroécologie (Duru et al., 2015). Cependant, l’AR semble trouver plus facilement un écho auprès d’une grande diversité d’acteurs (Paolini et al., 2024), à condition de définir des garde-fous pour éviter le greenwashing (Giller et al., 2021).
Pour accompagner cette transition et éviter les écueils, comme par exemple réduire l’AR au seul enjeu carbone (Lescuyer et al., soumis), plusieurs difficultés sont rencontrées. Côté recherche, les études concernant la biodiversité dans les sols et aérienne dans les parcelles et dans les paysages sont généralement conduites séparément, ce qui ne facilite pas la conception de pratiques favorisant conjointement ces différentes composantes. D’autre part, les évaluations de différents systèmes de culture agroécologiques sont souvent faites sur des systèmes figés (ex. essais à long terme avec la même rotation) alors que la résilience des systèmes repose sur leur capacité à évoluer et à s’adapter en fonction de l’évolution de leur environnement. Par ailleurs, les comparaisons sont souvent faites sur des systèmes aboutis et stabilisés (Duru et al., 2022), alors que dans la réalité beaucoup de sols sont dégradés (Panagos et al., 2024) et de nombreux paysages simplifiés (peu d’espèces cultivées, parcelles de grande taille). Ainsi, en France, dans plus de 85 % des exploitations agricoles, plus de 80 % de la sole cultivée ne sont occupés que par trois cultures au maximum (Vialatte et al., 2023).
Un deuxième type de difficultés provient du fait que les pratiques agroécologiques sont complexes à mettre en œuvre car dépendantes du contexte pédoclimatique et de l’histoire culturale des parcelles (Bullock et al., 2022). Ainsi, la variabilité dans les relations entre les diverses dimensions de la biodiversité et les services écosystémiques, n’autorise pas de recommandations standardisées en termes de pratiques mais simplement la formulation de principes. L’ACS notamment repose sur trois principes clefs : réduction du travail du sol, couverture permanente du sol, diversification végétale dans le temps (allongement des rotations) et dans l’espace (mélanges variétaux et association d’espèces). L’AB est basée sur quatre grands principes, de santé, d’écologie, d’équité et de précaution (IFOAM 2005) et de grands objectifs : interdiction des pesticides et engrais de synthèse, non utilisation d’OGM, obligation de rotation culturale, de fertilisation organique, de bien-être animal, de prévention des maladies et ravageurs, de respect de l’environnement. Elle se conforme à plusieurs cahiers des charges techniques cadrant pratiques et produits par des listes positives et négatives (cf. EU, 2018). Pour l’AR, ce sont souvent les mêmes types de principes que ceux de l’ACS et/ou de l’AB qui sont retenus (Sela et al., 2024). Ces principes sont des lignes directrices à garder à l’esprit lors du choix des pratiques et ne prescrivent aucune pratique spécifique ; le choix des pratiques dépendant du contexte. Ainsi, aucune pratique ne peut être qualifiée de « régénératrice » en soi, car son impact dépendra du contexte et des contraintes de l’agriculteur ainsi que du résultat des interactions avec les autres pratiques adoptées. En d'autres termes, il s'agit d'une « Trajectoire de progression », appelée ici régénération (Emmett, 2025). Une difficulté est qu’il peut apparaître des propriétés émergentes provenant de synergies ou d’effets rebonds qui facilitent l’atteinte des objectifs ou au contraire la freinent. Par exemple, l’arrêt du travail du sol, bénéfique pour la biodiversité de celui-ci, limite le contrôle des adventices, du moins dans un premier temps, ce qui entraine généralement un surcroît d’usage d’herbicides, et inversement l’arrêt des herbicides entraine une augmentation du travail du sol. Enfin, le risque est de privilégier certains enjeux plutôt que d’autres, le carbone plutôt que la biodiversité par exemple (Rehberger et al., 2023). Pour éviter le greenwashing, plusieurs recherches insistent à juste titre sur la nécessité de mettre en œuvre des approches basées sur les résultats et non pas sur les seuls moyens (Mambo et Lhermie, 2024). Un tel objectif montre le besoin, à l’échelle de l’exploitation, d’indicateurs d’état pour aider au pilotage des pratiques dans une diversité de contextes, voire le besoin, à l’échelle du paysage/territoire, d’indicateurs d’impact fonctionnel pour aider à la gouvernance (cf. méthode DPSIR – EEA, 2002 ; Duru et al., 2015).
L’objectif de cet article est donc de répondre à ces enjeux, en proposant les bases d’une stratégie pluriannuelle de régénération des agroécosystèmes via une séquence de pratiques à enclencher, adapter ou supprimer, de façon à maximiser les SE sous-tendant la production agricole tout en réduisant les impacts négatifs sur l’environnement. Dans une première partie, nous résumons les principales connaissances à mobiliser pour concevoir des systèmes agroécologiques. Puis, nous proposons une stratégie de régénération des agroécosystèmes et des indicateurs de suivi pour l’implémenter.
Principales connaissances pour la régénération des agroécosystèmes
Cadre d’analyse
Le maintien des conditions endogènes propices à la nutrition et à la santé des plantes repose sur deux principaux piliers. D’une part, une gestion assurant une alimentation hydrique et minérale optimale des plantes et des propriétés de sol "suppressives" des ravageurs et des maladies (Tamburini et al., 2020). D’autre part, une réduction des perturbations chimiques et mécaniques du sol et, plus généralement, du vivant, condition nécessaire à l’établissement de processus permettant la régulation biologique des bioagresseurs (Duru et al., 2015).
Les couverts végétaux gérés (rotation de cultures, intercultures, variétés, bandes enherbées, AF…) et les habitats semi-naturels génèrent dans les sols, les parcelles et les paysages, une biodiversité dite associée, qui arrive spontanément dans l’espace géré par l’agriculteur, mais qui est filtrée par ses pratiques (Power, 2010). Biodiversités, planifiée et associée, en interaction avec les pratiques, déterminent la dynamique de la MO du sol et des services écosystémiques à l’agriculture (ex. régulations biologiques) mais aussi à la société (ex. régulation du climat) (fig 1). A l’inverse, certaines pratiques agricoles comme les excès de travail du sol, d’utilisation des intrants de synthèse, de simplification des paysages, impactent négativement la biodiversité, surtout associée, et in fine les services qu’elle fournit à l’agriculture comme à la société (Therond et Duru, 2019).
Processus écologiques sous-jacents à la fourniture de services écosystémiques liés au sol
Le sol est potentiellement le support d’une grande diversité de micro-habitats et d’organismes de tailles très variables (de quelques microns à plusieurs décimètres). Il agit comme un accumulateur, un transformateur et un milieu de transfert pour l’eau, le carbone, l’azote, le phosphore, le soufre, les sels minéraux et les ions métalliques. Les processus physiques et chimiques qui contribuent à la santé du sol sont fortement liés aux activités des organismes du sol ainsi qu’à la structure et au fonctionnement des racines (Abbott et Manning, 2015).
Rôle de la biodiversité dans les cycles biogéochimiques, la nutrition et la protection des plantes
Les microorganismes participent à la nutrition des plantes, à la rétention et disponibilité des nutriments et à la stabilité structurale du sol (Adhikari et Hartemink, 2016). Ils jouent aussi un rôle important dans la séquestration du carbone (Domeignoz-Horta et al., 2024), essentiellement par le biais de la nécromasse microbienne, notamment dans les sols à forte abondance fongique (Khangura et al., 2023) et forte teneur en argile (Kirschbaum et al., 2020). Stocker du carbone dans le sol implique non seulement de mettre en œuvre des pratiques favorisant la biomasse microbienne, en particulier les bactéries solubilisatrices de minéraux et les champignons qui ont la capacité d’utiliser le carbone « liquide » provenant des exsudats racinaires, mais aussi de limiter le lessivage des particules fines du sol (argiles) en favorisant leur agrégation par des ‘colles biologiques’, i.e. des polysaccharides extracellulaires provenant de certaines bactéries et champignons, mycorhiziens (Hossain, 2021) comme non mycorhiziens (Stuart et al., 2024).
La diversité des microbes associés aux racines est également cruciale pour la santé des plantes. Celles-ci peuvent façonner le microbiome de la rhizosphère, notamment lors d'une attaque d'agents pathogènes ou d'invertébrés ravageurs. Elles peuvent alors recruter des microorganismes protecteurs spécifiques qui produisent des antibiotiques naturels ou encore des éliciteurs, renforçant la capacité des plantes à se défendre contre les bioagresseurs (Berendsen et al., 2012). Les vers de terre jouent aussi un rôle particulièrement important dans la structuration du sol (porosité) et dans la décomposition des MO mais aussi la croissance (Van Groenigen et al., 2014) et la santé des plantes (Meghvansi et al., 2011).
Les organismes du sol utilisent comme principale ressource énergétique les résidus de culture et les rhizodépôts. Parmi ces derniers, les exsudats racinaires représentent un « carburant » rapidement et facilement assimilé par de nombreux organismes dont ils stimulent le développement et la prolifération tout autour de la racine. En retour, les microorganismes (bactéries, champignons) stimulent l’exsudation (Lebeis et al., 2015) et élicitent des réactions de la plante-hôte (Pieterse et al,. 2014) permettant le développement de véritables chaînes alimentaires. Les bactéries (plus rapides à se multiplier) interviennent les premières puis arrivent les champignons (Jacoby et al., 2017), les consommateurs primaires tels les protozoaires et vers de terre, et enfin les consommateurs secondaires comme les collemboles et acariens. Tous ces organismes contribuent aux processus d’agrégation des sols et donc à leur structuration, qui, en retour, détermine les premiers. Ainsi, dans un sol compacté, caractérisé par une diminution de la diffusion de l’oxygène et de la diversité des micro-niches trophiques et physicochimiques, la richesse et la diversité spécifiques des communautés microbiennes sont moins élevées qu’en sol bien structuré (Fierer et Jackson, 2006), et les bactéries oligotrophes, adaptées au stress car à stratégie écologique stable et lente de type K, dominent (Peng et al. 2022). Inversement, en sol bien structuré (davantage de ressources et donc de micro-niches disponibles, favorisant des communautés plus riches et diversifiées – op. cit.), la plante bénéficie de la présence de bactéries copiotrophes à stratégie essentiellement ‘explosive’ de type ‘r’, opportunistes i.e. capables, sous l’effet des exsudats racinaires, de rapidement se multiplier et d’exploiter, à leur profit et celui de la plante, les ressources présentes dans le sol (Peng et al., 2022). Ces états sont conformes aux travaux de Dini-Andreote et al. (2015), qui montrent que la succession écologique des communautés microbiennes tend à favoriser une sélection plus déterministe et moins stochastique à mesure que le milieu devient plus structuré et stable, par exemple lorsqu’un sol initialement compacté développe peu à peu une meilleure structure, plus riche en niches écologiques et donc plus propice à la diversité microbienne. La diversité des cultures tend à renforcer la diversité et par-là les fonctions des organismes du sol, donc à renforcer les premières (Wang et al., 2022).
L’homéostasie pH-Eh du sol et des plantes, clé de voûte de processus écologiques majeurs
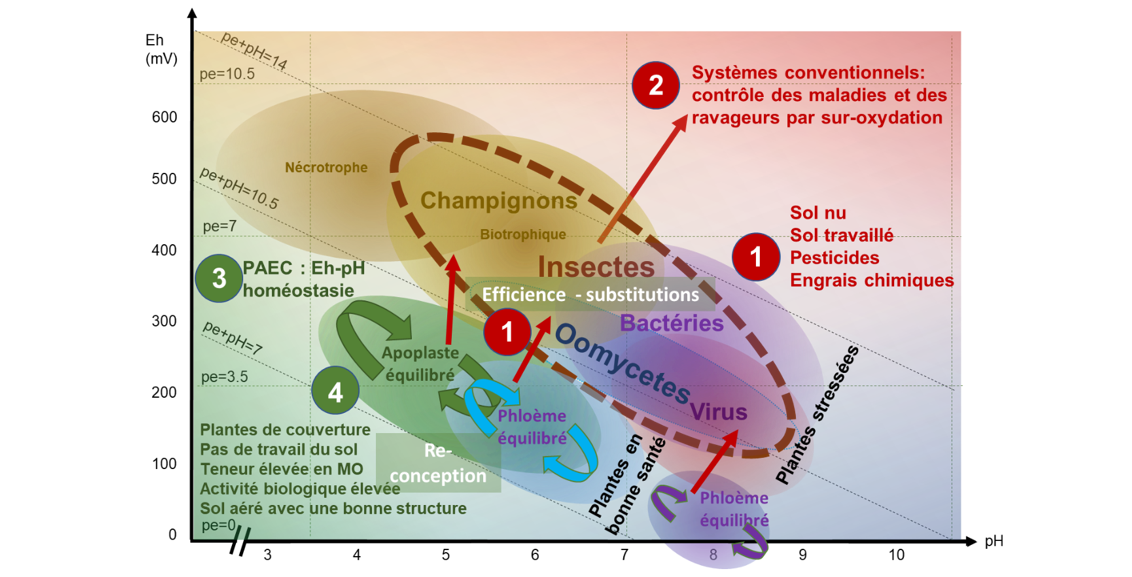
La revue de littérature de Husson et al. (2021) montre l’importance du potentiel redox (Eh) et du pH sur la santé des sols et des plantes, et inversement. Ainsi, les différents types de ravageurs et pathogènes se développent dans des niches Eh-pH spécifiques (fig. 2). Ces conditions Eh-pH de la plante sont fonction du génotype et des conditions de culture, sachant que tout stress biotique ou abiotique conduit à une oxydation (Husson et al., 2021).
Alors que la protection des plantes consiste généralement à détruire les bioagresseurs par suroxydation via l’application de pesticides, la protection agroécologique des cultures (PAEC) correspond à une stratégie alternative reposant sur le maintien dynamique de l’homéostasie Eh-pH dans le sol et dans les plantes. La gestion du sol nécessaire à la mise en place de la PAEC nécessite de repenser les systèmes de culture afin d’optimiser la production photosynthétique et surtout de fournir, grâce à elle, l’énergie nécessaire à la création et au maintien d’une structure endogène optimale du sol, via une augmentation de la MO du sol et une augmentation de l’activité et de la diversité de ses communautés biologiques (fig 2). Cette structure favorise la sélection par les plantes de microorganismes favorables en permettant la circulation et le stockage de l’eau tout en assurant la circulation de l’air, en créant une diversité de niches écologiques et en en limitant leurs fluctuations Eh-pH (Husson et al., 2021 ; Husson, 2023).
Ainsi, les pratiques telles que le sol nu, le travail du sol, l’usage de pesticides et d’engrais minéraux et une faible diversité des cultures, conduisent à une oxydation du sol et des plantes qui deviennent, de ce fait, sensibles aux bioagresseurs (La Scala et al., 2008 ; Modak et al., 2019 ; Rasmusson et al., 2020 ; Virk et al., 2024). Les systèmes reposant sur la biodiversité planifiée (cultures et couverts végétaux), des apports d’intrants de synthèse réduits et bien ciblés (ex. apports foliaires d’oligo-éléments ; Noreen et al., 2018), et sur une réduction du passage d’engins lourds et du travail du sol pour mieux bénéficier d’un retour accru de résidus au sol, créent des conditions optimales en termes d’équilibre pH-Eh, de richesse en MO et de structure endogène (générée par l’activité biologique) (Husson et al., 2016, Husson et al., 2021 ; Sher et al., 2024). Ces équilibres électro-chimiques sont alors défavorables aux bioagresseurs (valeurs Eh trop basses) et au contraire favorables, via la richesse en MO et une bonne structure endogène, à une activité biologique intense (Bender et al., 2016), réduisant la capacité des pathogènes à coloniser le sol (Van Elsas et al., 2012). Les sols deviennent alors « suppressifs », c’est à dire défavorables aux bioagresseurs (Bonanomi et al., 2018 ; Löbmann et al., 2016), et fournissent d’autres services à l’agriculture voire à la société.
Cette approche homéostatique Eh-pH de la santé des sols et des plantes offre une perspective originale pour démêler les nombreuses interactions Génotype x Environnement x Pratiques x Ravageurs/Pathogènes et ainsi clarifier l’impact des pratiques culturales sur la santé des sols et des plantes.
Les études sur la nutrition des plantes ont souvent ignoré le rôle important des organismes saprophages, intervenant dans la décomposition des résidus organiques et dans le cycle des éléments nutritifs. En effet, pour véritablement piloter les cycles du carbone et des éléments nutritifs via les pratiques culturales, il faut tenir compte du fait que les communautés hétérotrophes dans leur ensemble (saprophages, phytophages/herbivores, prédateurs/carnivores), maillons essentiels de ces cycles, sont limitées par les ressources carbonées riches en énergie (Soong et al., 2020). En conséquence, la gestion régénératrice des sols consiste à conserver voire amplifier les pools de nutriments auxquels les plantes peuvent accéder pour élaborer leur biomasse (ressources carbonées). En outre, elle vise à améliorer l’efficience de la transformation de cette biomasse morte, appelée MO fraiche (résidus de cultures et couverts, produits résiduaires organiques) en MO dite évoluée (biopolymères, libres ou associés aux particules minérales). Ces éléments nous permettent de synthétiser les effets attendus de différentes pratiques agricoles sur les propriétés physicochimiques et biologiques du sol. Les données du tableau 1 montrent des effets positifs des couverts végétaux et rotations de culture, et des effets plutôt négatifs du travail du sol et des intrants de synthèse, sauf pour la régulation des bioagresseurs, sachant que souvent ces pratiques limitent voire empêchent, à terme, l’expression des SE que peut rendre la biodiversité. Ces effets et leur importance varient selon les conditions, en particulier en fonction du climat et des systèmes de culture.
Tableau 1. Effets des leviers agronomiques sur les propriétés physicochimiques et biologiques d’un sol, et services associés. Noter qu’en situation de sol dégradé, certaines pratiques agricoles intensives, aux effets négatifs sur les propriétés d’un sol en bon état, sont souvent nécessaires à la production de biomasse végétale, étape initiale incontournable de la régénération des sols.
Légende des symboles : + et -, effets respectivement positifs et négatifs des pratiques agricoles sur les caractéristiques du sol ; le nombre de + et de – indique l’intensité de l’effet ; / signifie positif ou négatif selon les situations
Pratiques agricoles | Propriétés du sol | Effets pour sols dégradés | Effets pour sols restaurés | |||
| Homéostasie Eh-pH Husson, 2013 ; Husson et al., 2016 ; Husson et al., 2018 ; Husson et al, 2021 | Biodiversité
de Graaf et al, 2019 McLaughlin & Mineau, 1995 ; Beaumelle et al., 2023 ; Lupwayi et al., 2024 | Structuration endogène du sol et contrôle de l’érosion Bronick & Lal, 2005 ; Howe et a.l, 2024; Adetunji et al., 2020 ; Christel et al, 2021 | Régulation des bioagresseurs Deguine et al, 2023 ; Datnoff et al, 2007 ; Derrouch et al, 2024 ; Sarthou et al., ; 2024a ; Sarthou et al, 2 024b | Structuration endogène du sol et contrôle de l’érosion Jeyaseelan et al., 2024 ; Zhang et al, 2022 ; Mehra et al, 2018 ; Prashar & Shah, 2016 ; Wortmann et Dang, 2020 | Régulation des bioagresseurs Deguine et al., 2023 ; Sias et al., 2021 ; Jastoria et al., 2023 ; Derrouch et al., 2024 ; Sarthou et al., 2024a ; Sarthou et al., 2024b |
Labour ou travail du sol profond | -- | -- | -- (moyen terme) + (court terme, fissuration sur sol compacté qui permet l’installation des cultures/couverts) | + (adventices, ravageurs)
| -- | - (antagonistes d’agents phyto-pathogènes) |
Travail du sol superficiel | - | - | - | + (adventices) | - | + (adventices) |
Passage d’engins lourds (récolte et charrois) | - - | - - | - - | - | - | / |
Drainage | + | + | + | + | / | / |
Fertilisation minérale | - | - | + (via amélioration de la photosynthèse et du stockage de MO) | |
|
|
Correction des déséquilibres nutritionnels (dont chaulage) | + | + | / | / puis + via amélioration physio / syst. immunitaire des plantes | / et + à terme via accroissement des résidus | / puis + via amélioration physio / syst. immunitaire des plantes |
Amendements et apports organiques | + | + | + | + | + | + |
Fongicides et insecticides | - | - - | /(+ via production de biomasse) | + à court terme
| / puis - via nuisibilité pour communautés de champignons mycorhiziens | - à moyen et long termes via nuisibilité pour antagonistes de bioagresseurs |
Herbicides | - | - | / | + à court terme
| / | - via apparition résistances et nuisibilité pour auxiliaires granivores |
Rotation/ couvert | + | + | + | + | + | + |
Processus écologiques dépendant de la diversité des plantes dans l’espace et dans le temps
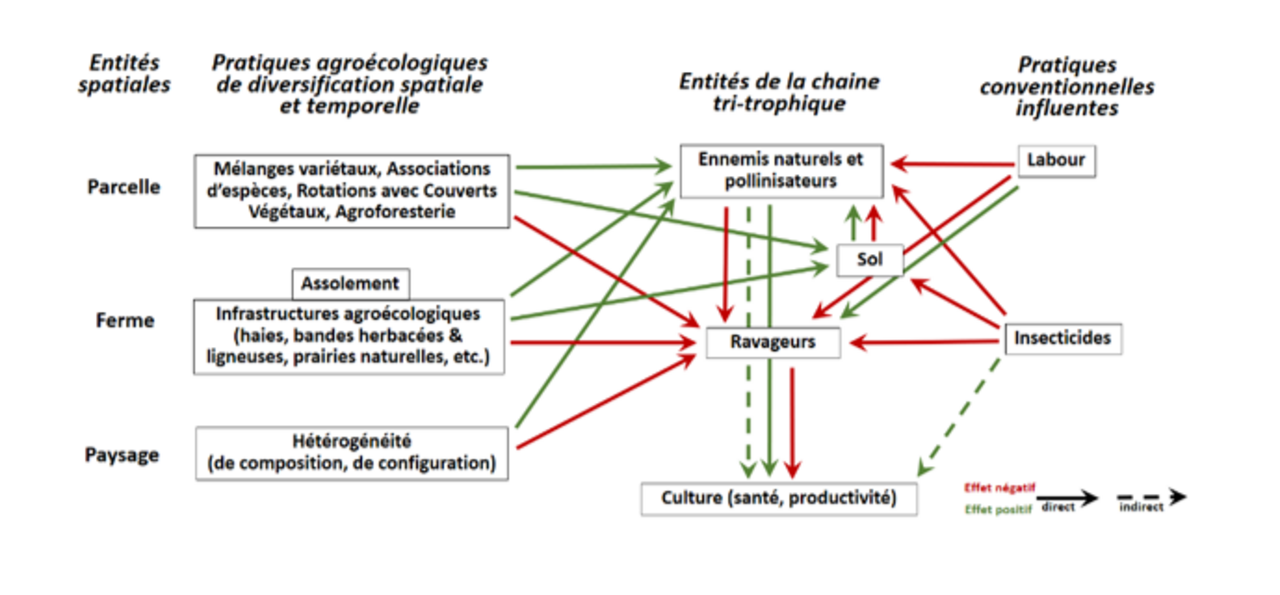
Les effets de la diversification végétale cultivée dans l’espace (mélanges variétaux et d’espèces herbacées et ligneuses, assolement) et dans le temps (rotation), mais aussi des habitats semi-naturels (haies, bandes fleuries, bandes culturales extensives) sur la dynamique des ravageurs des cultures et de leurs ennemis naturels, font l’objet de nombreuses recherches. Loin d’être univoques, les résultats affichent néanmoins des tendances claires : Holmes et Blubaugh (2023), dans une méta-analyse de 2nd ordre portant sur des travaux publiés sur 30 ans, montrent que si la diversité des herbivores augmente avec la diversité végétale, l’abondance de ceux-ci et surtout les dommages qu’ils causent aux cultures, diminuent. Ces tendances générales (fig. 3) masquent des réponses plus nuancées selon les espèces de plantes et d’insectes : les insectes phytophages spécialisés réagissent négativement à la diversité végétale, alors que les généralistes y répondent de façon neutre ou positive. En moyenne, la diversification végétale favorise la diversité fonctionnelle des pollinisateurs mais aussi des ennemis naturels des ravageurs, aux dépens de ces derniers et de leurs dommages (Letourneau et al., 2011 ; Dainese et al., 2019). Ainsi, jusqu’à 50 % des effets négatifs de la simplification des paysages sur la pollinisation et le contrôle biologique des ravageurs, sont dus à des pertes de richesse spécifique des pollinisateurs et ennemis naturels, avec des conséquences négatives sur le rendement des cultures, particulièrement du fait d’un déficit de pollinisation (Dainese et al., 2019).
Des travaux récents prouvent que la configuration spatiale du paysage (taille des parcelles et répartition des cultures) a un effet au moins aussi important que celui de sa composition (diversité des espèces cultivées et semi-naturelles) (Vialatte et al., 2023). D’autres travaux ont montré que les pratiques à l’échelle de la parcelle influencent aussi la dynamique du contrôle biologique des ravageurs (Karp et al., 2018 ; Tamburini et al., 2019). Ils montrent, par exemple, que l’ACS favorise les prédateurs généralistes du sol (carabes, araignées, staphylins) mais aussi les prédateurs aphidiphages aériens, et in fine favorise la régulation des pucerons par prédation et parasitisme au point, dans leur cas d’étude, de pouvoir compenser les effets négatifs de la simplification des paysages. Enfin, selon Chabert et Sarthou (2017), l’ancienneté du dernier labour l’emporte sur le type de système (ACS, AB, AF) et explique mieux le service de régulation naturelle des pucerons par une espèce de syrphe, que la complexité du paysage. La prééminence, pour certains groupes biologiques tout au moins, des pratiques agricoles intra-parcellaires sur le paysage, pourrait expliquer l’absence d’effets de la complexité paysagère sur la diversité des micro-hyménoptères parasitoïdes et des coléoptères prédateurs (Carabidae et Staphylinidae), observée par Shackelford et al. (2013) dans une méta-analyse portant sur 46 études. Dans la même logique, la méta-analyse de 2nd ordre (98 méta-analyses et environ 42 000 comparaisons) de Tamburini et al. (2020) sur l’effet des pratiques agroécologiques pour le contrôle biologique, montre que la diversification végétale cultivée est le levier le plus fort parmi les 6 testés, suivie de l’arrêt de l’usage des pesticides puis de la diversification végétale non cultivée. Concernant l’importance de cette dernière, Crowther (2023) ont démontré dans une méta-analyse portant sur 40 études, que les bordures de champs fleuris, les naturelles davantage que les semées, augmentent significativement l’abondance et la diversité des ennemis naturels des ravageurs des cultures, et réduisent significativement l’abondance de ces derniers mais aussi leurs dommages dans les cultures. Une manière très convaincante de démontrer l’impact sur la dynamique des ravageurs de la présence de leurs ennemis naturels, est d’exclure ces derniers de certaines zones des cultures et de voir comment évoluent les ravageurs par rapport aux zones témoins, i.e. auxquelles prédateurs et parasitoïdes accèdent toujours. Boldorini et al. (2024) ont réalisé une méta-analyse de 86 études de ce type (soit 317 comparaisons), et observent qu’en moyenne les ravageurs augmentent de 270% dans les zones d’exclusion, et que la présence d’auxiliaires augmente les rendements de 25% dans les zones témoins.
Tamburini et al. (2020) mettent aussi en évidence que les deux leviers les plus importants pour soutenir la pollinisation, sont la diversification végétale non cultivée (diversité constitutive des habitats semi-naturels ou infrastructures agroécologiques) puis l’arrêt de l’usage des pesticides. Enfin, une récente synthèse propose de classer différents leviers quant à leurs effets sur la régulation des bioagresseurs, la séquestration du carbone et les fonctions régulatrices du sol vis-à-vis de l’eau (Vialatte et al., 2023). Les auteurs ont jugé, ‘à dire d’experts’, que trois leviers agroécologiques sont d’une efficacité notable pour favoriser ces services, et sont, par ordre d’effet décroissant : l’agroforesterie, la rotation des cultures et les couverts végétaux, les cultures associées. Notons toutefois, concernant l’agroforesterie et ses effets sur le service de contrôle biologique des ravageurs des cultures, qu’une méta-analyse récente conclut sur son potentiel effectivement prometteur en régions tempérées (en régions tropicales, ses effets positifs pour ce service sont largement démontrés), mais aussi sur le caractère encore peu représentatif des 12 études seulement ayant servi de base à ce travail (Staton et al., 2019).
Enfin, concernant les régulations microbiologiques influençant la dynamique des agents phytopathogènes, la diversification des cultures, notamment par l’allongement des rotations, les cultures associées ou les couverts végétaux, favorise l’augmentation de la diversité microbienne des sols, en particulier rhizosphérique. Cette diversification accroît la probabilité de présence de micro-organismes antagonistes des agents phytopathogènes, contribuant ainsi au développement de sols suppressifs (Jayaraman et al., 2021). Cet effet est renforcé dans les systèmes à perturbation mécanique réduite et avec présence continue de résidus organiques en surface, systèmes qui créent un environnement favorable à l’établissement et au maintien de communautés microbiennes bénéfiques. Toutefois, la construction de la suppressivité des sols reste progressive et dépend des interactions spécifiques entre pratiques culturales, conditions édaphiques et pathosystèmes ciblés (Sarthou et al., 2024c).
Comment régénérer les agroécosystèmes dégradés et/ou trop simplifiés
Stratégie globale
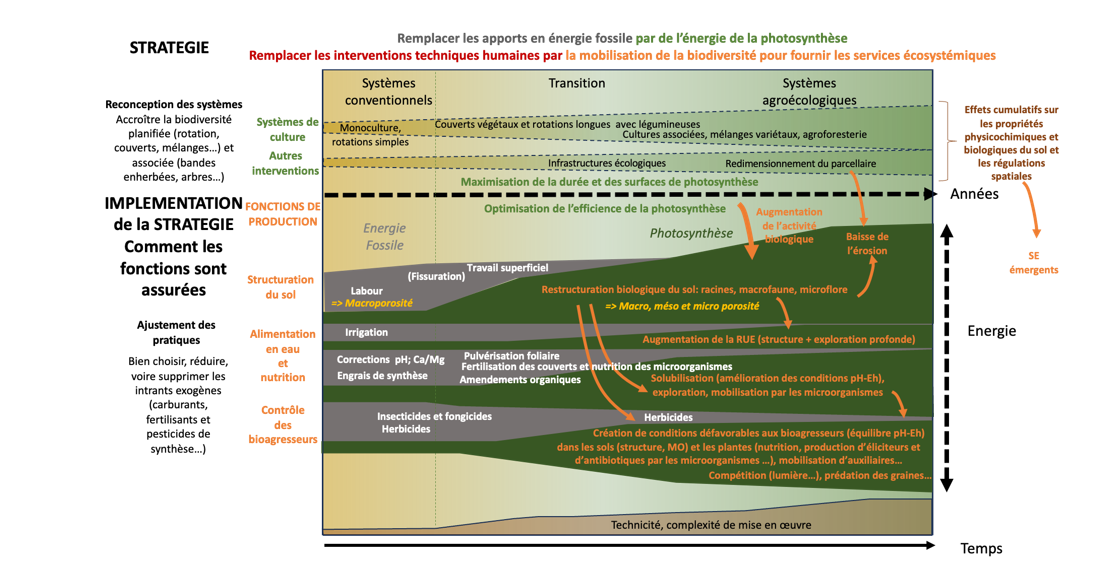
Le principe de base de la régénération des agroécosystèmes fondés sur le modèle industriel est de remplacer autant que possible l’énergie culturale dite industrielle, provenant directement ou indirectement d’une énergie fossile, par l’énergie culturale dite biologique, issue directement ou indirectement de la photosynthèse (Gliessman et al., 2022). Les pratiques d’allongement des rotations, incluant l’implantation de couverts végétaux pendant les intercultures (d’au minimum 6-7 semaines), de cultures associées voire de plantes compagnes, de prairies temporaires, la mise en place d’infrastructures agroécologiques pérennes (haies, talus, prairies naturelles…) ou temporaires (bandes culturales extensives, bandes enherbées et bandes fleuries…), la réduction voire l’arrêt, au moins séquentiel, du travail du sol, permettent d’optimiser spatialement et temporellement, quantitativement et qualitativement, la production végétale et ses co-produits (biomasse verte, pollen, nectar floral et extra-floral, domaties pour les auxiliaires, nécromasse en fin de cycle). Par effet de cascades trophiques, la diversification des cultures permet d’optimiser les processus écologiques générés par la biodiversité qui en dépend, et finalement les services écosystémiques attendus i.e. l’énergie culturale biologique qui viendra remplacer tout ou partie de l’énergie culturale industrielle (Fig 4).
La première étape nécessite un diagnostic basé sur des analyses de terre (par ex. Bucaille, 2023) complétées par la comparaison du niveau d’intrants par rapport à des référentiels régionaux de systèmes de culture performants (e.g. IDEA 4 – Zahm et al., 2020).
Un premier point clef est de s’assurer que le niveau de la photosynthèse est maximal, en l’améliorant si nécessaire par la fertilisation, y compris foliaire ; en particulier pour lever des carences induites par des conditions Eh-pH du sol qui ne peuvent être levées rapidement via la régénération du sol. En effet, si des blocages par excès ou carences de minéraux ne sont pas d’abord levés, l’agroécosystème ne pourra pas produire assez de biomasse, et les effets attendus de la biodiversité ne pourront pas complètement s’exprimer. Il peut s’agir par exemple de corriger les déséquilibres cruciaux (pH, Ca/Mg, K/Mg[1] ; valeurs indicatives à viser, respectivement : 6,5 à 7,5 ; 3,5 à 5,5 ; 0,2 à 0,4), parfois simplement les excès ou les manques d’eau ou de porosité (donc d’air) dans les sols. Des amendements calciques ou magnésiens peuvent alors être réalisés (chaux, calcaire, dolomie) pour augmenter le pH des sols acides, en les combinant si possible à des amendements organiques car la MO du sol tend elle-aussi à limiter les valeurs extrêmes de pH et de Eh (Rowley et al., 2018). Elle contribuera également à conférer une structure aérée, dite grumeleuse, et stable aux agrégats du sol par le biais des communautés microbiennes auxquelles elle servira de substrat (Rabbi et al., 2020).
Le deuxième point clef est en effet d’utiliser au mieux cette énergie culturale biologique pour (re)créer des conditions favorables aux cultures en améliorant la structure du sol afin de fournir le gite et le couvert à une armée « d’ouvriers spécialisés » qui vont soutenir les services de restructuration du sol (macro-, méso- et microfaune + microflore + plantes), de « digestion » des éléments nutritifs (solubilisation, chélation), de production d’antibiotiques naturels etc. (Lavelle et al., 2020 ; Wang et al., 2024 ; Devi et al., 2022 ; Tyc et al., 2014). La structure du sol sera plus rapidement améliorée par les organismes telluriques si les particules fines (argiles minéralogiques, limons fins) s’agrègent aisément entre elles et avec les MO, en décomposition et évoluées.
Ainsi, c’est l’optimisation de l’utilisation de l’énergie solaire incidente qui est améliorée par des plantes photosynthétiquement très actives, mais aussi l’efficience de transformation des résidus végétaux en MO évoluée, véritable condensé en minéraux dont les plantes et les organismes du sol ont besoin pour se développer. En outre, le stockage du carbone dans les sols est corrélé positivement à l’efficience de l’utilisation du carbone par les microorganismes du sol (Bertrand et al., 2019), efficience qui dépend notamment de la disponibilité en éléments minéraux (N, P, S…), déterminée par les équilibres stœchiométriques de la MO des sols, soit C:N :P :S = 10000 :833 :200 :143 (Kirkby et al., 2013, 2014). Respecter cet équilibre stœchiométrique est un facteur clé pour améliorer le stockage de carbone, au moins 4 fois plus important que d’autres facteurs comme les apports de carbone (Tao et al., 2023 ; Encadré 1).
Leviers agroécologiques pour restaurer la fertilité, la diversité et les régulations naturelles
Les principaux leviers de la régénération des agroécosystèmes sont désormais bien identifiés : promotion de la biodiversité planifiée et gestion raisonnée des intrants pour promouvoir et ne pas remettre en cause cette première. Ils visent à réduire les perturbations physiques et chimiques du sol, maintenir une couverture végétale permanente, préserver des racines vivantes tout au long de l’année, accroître la diversité végétale. Leur mise en œuvre exige une combinaison progressive de leviers directs et indirects dans le temps, en commençant par lever les facteurs limitants majeurs pour tirer parti des synergies et éviter les effets rebonds, par exemple un besoin accru d’intrants de synthèse.
Diagnostic de l’agroécosystème et priorités d’action
Lorsqu’un sol est fortement dégradé, l’action prioritaire consiste à améliorer sa structure par un apport de carbone, éventuellement un amendement calcique ou magnésien (Bucaille, 2023) et à maintenir un minimum de travail du sol, parfois profond mais sans retournement (passage d’un fissurateur voire d’un décompacteur), afin de maintenir une macroporosité minimale permettant l’implantation des cultures et des couverts (Sarthou et al., 2024a). L’implantation de couverts végétaux durant les intercultures améliore également la porosité (de nature biologique, donc plus stable), la stabilité structurale et enrichit le sol en matière organique, à condition que les déficits éventuels en calcium, se traduisant par une insuffisante floculation des argiles, soient préalablement corrigés par des apports de CaCO3 (calcaire brut, pouvant être transformé en chaux) si le pH est trop acide (pH < ~6,5), ou de CaSO4 (gypse) s’il ne doit pas être corrigé (pH entre ~6,5 et ~7,5). Par ailleurs, la diversification végétale dans le temps (rotations allongées) et dans l’espace (plantes compagnes, mélanges variétaux et d’espèces) maximise la couverture végétale et stimule la photosynthèse, ce qui in fine améliore l’équilibre Eh-pH via les effets structurants des systèmes racinaires et le pouvoir tampon sur le couple pH-Eh de la MO du sol (Husson et al., 2016).
Vers une fertilité de plus en plus endogène des sols
L’allongement des rotations, incluant cultures d’hiver, couverts intermédiaires, cultures de printemps et légumineuses, en plus de la limitation du travail du sol (dès qu’elle est possible), améliore la nutrition végétale tout en réduisant le recours aux engrais azotés de synthèse, limite les bioagresseurs, et finalement contribue significativement à l’amélioration de la fertilité endogène des sols (Smith et al., 2008). Par ailleurs, les légumineuses jouent un rôle clef dans les régulations redox via la production d’H2 par la nitrogénase, gaz qui soutient indirectement la croissance et la santé des plantes via ses effets sur les micro-organismes du sol (Azotobacter, Azomonas, etc.) (Islam et al., 2023 ; Zulfiqar et al., 2021). Les légumineuses constituent donc un levier de premier plan pour la réduction des intrants de synthèse (engrais et pesticides), et sont très complémentaires des amendements organiques pour la mise en place d’une bonne fertilité endogène des sols cultivés (Johnston et Poulton, 2018), prérequis à une plus grande autonomie vis-à-vis des engrais de synthèse. Notons que durant la phase de régénération des sols et tant que la richesse optimale en éléments minéraux du sol n’est pas acquise, deux pratiques complémentaires sont à conseiller : (i) pulvérisations foliaires d’azote réduit, d’acides fulviques et autres biopolymères avec des macro- et oligo-éléments sur les plantes en développement pour corriger rapidement les carences et leurs effets sur les rendements, et (ii) apport d’éléments minéraux sur les résidus de culture à forts C/N, afin d’améliorer leur incorporation dans la MO des sols (Sarthou et al., 2024a ; et voir encadré 2). Au fil du temps, ces deux pratiques sont de plus en plus substituables jusqu’à ce qu’elles deviennent superflues, car peu à peu et parallèlement : (i) le fonctionnement du sol s’améliore (minéralisation de la MO), limitant le besoin de fertilisation foliaire, et (ii) le stock d’éléments minéraux augmente, améliorant le coefficient isohumique k1 et donc réduisant la nécessité d’en apporter sur les résidus. Toutes deux accélèrent la cinétique d’amélioration du statut organique des sols (et donc du stockage de carbone) et celle du rendement des cultures et de leur résilience.
De plus, l’amélioration progressive de la structure du sol, de la disponibilité des minéraux et des différentes formes de MO du sol impacte les populations de microorganismes, favorisant des populations à forte efficience de l’utilisation du carbone et accélérant ainsi la régénération des sols (encadré 3).
Vers des systèmes de culture de plus en plus résistants aux bioagresseurs
La baisse de l’usage des fongicides et insecticides de synthèse dans les systèmes agricoles (non AB, car les systèmes AB n’y ont par définition pas recours) ayant régénéré leurs sols et leurs infrastructures agroécologiques, n’est ni systématique ni flagrante. Cependant de plus en plus de travaux de recherche démontrent une fréquente moindre dépendance des systèmes ACS aux insecticides (e.g. Basch et al., 2015 ; Kesavan et Malarvannan, 2010 ; Lefèvre, 2024), aux fongicides (e.g. Basch et al., 2015 ; Kutcher et al., 2011 ; Lefèvre, 2024), et même un potentiel, dans des systèmes avec très bonne fertilité endogène des sols et pratique des couverts végétaux, de moindre utilisation d’herbicides en systèmes ACS par rapport à des équivalents en conventionnel (Anderson 2005, 2008 ; Derksen et al., 1996 ; Lefèvre, 2024). Les meilleures régulations d’agents phytopathogènes et de ravageurs en systèmes avec sol régénéré, sont en effet permises par un meilleur fonctionnement du sol (cf 2.2.2.) en tant qu’habitat d’espèces auxiliaires (prédateurs et parasitoïdes des ravageurs des cultures, et bactéries et champignons antagonistes des agents phytopathogènes – Sarthou et al., 2024b, 2024c). La mise en place d’infrastructures écologiques (haies, bandes enherbées) et plus globalement le renforcement de la diversité paysagère (dont celle des cultures), soutiennent eux aussi les auxiliaires et limitent l’impact des ravageurs dans les cultures (voir paragraphe 2.3).
S’inscrire dans le temps long
Sur le temps long, deux leviers porteurs d’impacts potentiellement forts sur les performances agronomiques et environnementales, méritent d’être mis en œuvre en parallèle : d’une part l’agroforesterie, pour ses effets sur le microclimat, la régulation biologique, l’érosion et l’infiltration de l’eau (ex. : plantations en keylines) ; et d’autre part le redimensionnement parcellaire et la coordination territoriale des assolements, afin d’améliorer l’influence positive de la composition et configuration du paysage sur les régulations biologiques et donc la pression des bioagresseurs et la pollinisation.
Suivre la trajectoire de régénération
Enfin, le suivi de la régénération de l’agroécosystème et de ses compartiments sol cultivé, infrastructures agroécologiques, biodiversités planifiée et associée, eau et hétérogénéité spatiale, résultantes multidimensionnelles des choix de systèmes et de pratiques en interaction avec les facteurs naturels, doit être engagé rapidement. L’objectif est de déterminer l’état du système avant les changements liés à la transition (‘point zéro’), et de réaliser des points d’étape réguliers à l’aide d’indicateurs fiables de pression (i.e. de moyens) mais préférentiellement d’état (i.e. de résultats) voire d’impact sur les fonctionnalités (cf. méthode DPSIR – EEA, 2002). Cela permet d’apporter à l’agriculteur une vision d’ensemble mais aussi détaillée et opérationnelle, lui permettant de suivre l’évolution des caractéristiques de son système au fur et à mesure de la mise en œuvre de ses pratiques. Par exemple, le ratio MO%/argile%, même s’il fait l’objet de discussion et n’est adapté que dans certaines limites (Poeplau et Don, 2023 ; Rabot et al, 2024 ; Sauzet et al., 2024) permet une évaluation rapide et simple de la fertilité physique, plus précisément de la sensibilité du sol à perdre sa structure, caractéristique sous influence à la fois de l’intensité des perturbations qui lui sont appliquées et de sa teneur en MO, partiellement dépendante du facteur précédent (Johannes et al., 2017). D’autres indicateurs, comme un simple test bêche et tout un référentiel d’indicateurs peuvent aussi être utilisés, comme proposés par Cousin et al. (2024). L’approche Eh-pH, plus technique, est utile pour ajuster les pratiques durant la phase de transition vers des agroécosystèmes matures (fig. 4). D’un point de vue opérationnel, l’indice de régénération, proposé par le mouvement Pour une Agriculture Du Vivant (PADV), analyse les pratiques agroécologiques mises en œuvre pour mesurer leurs impacts sur la santé des sols, des plantes et des paysages, identifier des axes de progrès, et valoriser les efforts des agriculteurs engagés dans la transition.
Conclusion : pour une agriculture véritablement régénératrice
Face aux enjeux sanitaires et environnementaux, développer des systèmes agroécologiques fondés sur les services écosystémiques à l’agriculture et minimisant les impacts environnementaux est une nécessité. L’articulation des recherches récentes sur les effets de la diversification végétale et des pratiques agricoles, tant sur les régulations biologiques que les cycles biogéochimiques, fournit un cadre permettant de définir des principes d’action pour renforcer la biodiversité sous-tendant ces services ainsi que sur l’abandon ou la réduction de pratiques ayant des effets adverses sur celle-ci. Cependant, une telle perspective présente des difficultés et des risques. Elle demande aussi un effort de formation, d’accompagnement (en particulier d’assurer la prise de risque) et de conseil.
Passer des principes à l’action est complexe car toutes les pratiques soutenant les services ne sont pas à mettre en place au même moment. Leur choix est dépendant du contexte : état de dégradation de l’agroécosystème, type de sol, de culture, de climat…. Aussi, nous avons défini des principes de régénération des agroécosystèmes permettant d’ordonnancer les pratiques à mettre en œuvre en fonction de la situation de départ. Et cela vaut pour toutes les formes d’agriculture se référant à l’agroécologie. Le seul cahier des charges en agriculture biologique, ou la simple mise en œuvre des principes en agriculture de conservation des sols, ne suffisent pas. Pour se maintenir dans une direction de progression, autrement dit sur une trajectoire de régénération de la biodiversité, il importe de piloter l’agroécosystème de manière agile, ce qui suppose de s’adapter chemin faisant avec l’aide d’indicateurs de pilotage. De tels indicateurs aideront à décliner les principes de la régénération des agroécosystèmes en pratiques de gestion. Cette déclinaison nécessite d’hybrider les savoirs scientifiques des chercheurs et empiriques des acteurs de terrain pour une bonne adaptation des pratiques à la diversité des contextes pédoclimatiques.
Régénérer les agroécosystèmes n’exclut pas l’utilisation des technologies génétiques, robotiques, numériques. Il importe cependant que ces technologies soient adaptées au besoin de pilotage des trajectoires de régénération. Autrement dit, les technologies doivent réellement être pertinentes pour accompagner la mise en œuvre des principes de l’agroécologie et non s’imposer à elle.
En agriculture régénératrice, tout comme en agriculture de conservation, qui reposent toutes deux sur des principes, le risque est d’aboutir à un choix de pratiques et d’objectifs d’évolution « à la carte », portés, par exemple, par un effet d’aubaine. Par exemple, les progrès peuvent être importants sur le cycle du carbone, mais moins bons voire nuls sur la biodiversité, du fait par exemple d’apports d’intrants de synthèse, notamment de pesticides, inadaptés. Ce risque est d’autant plus grand que le levier portant sur la diversification des cultures est faiblement mobilisé dans les cahiers des charges des promoteurs de l’AR (Lescuyer et al., soumis) où l’obligation stricte de non travail du sol est érigée en dogme. Pour éviter ce type de dérapage, les indicateurs de moyen (bilan azote, IFT) peuvent être mobilisés.
Sous cette réserve, le concept de régénération présente l’intérêt de faciliter le dialogue interdisciplinaire et inter-institutionnel pour aboutir à des solutions tenant compte des spécificités territoriales (Paolini et al., 2024).
Références bibliographiques
Abbott, L. K., & Manning, D. A. (2015). Soil health and related ecosystem services in organic agriculture. Sustainable Agriculture Research, 4(526-2016-37946).
Adetunji, A. T., Ncube, B., Mulidzi, R. & Lewu, F. B. (2020). Management impact and benefit of cover crops on soil quality: A review. Soil and Tillage Research 204, 104717.
Adhikari, K., & Hartemink, A. E. (2016). Linking soils to ecosystem services - A global review. Geoderma, 262, 101–111.
Anderson, R. L. (2005). A Multi-Tactic Approach to Manage Weed Population Dynamics in Crop Rotations. Agronomy Journal, 97, 1579‑1583.
Anderson, R. L. (2008). Diversity and No-Till: Keys for Pest Management in the U.S. Great Plains. Weed Science, 56 (1), 141‑145. https://doi.org/10.1614/WS-07-007.1
Angst, G., Mueller, K. E., Castellano, M. J., Vogel, C., Wiesmeier, M., & Mueller, C. W. (2023). Unlocking complex soil systems as carbon sinks: multi-pool management as the key. Nature Communications, 14(1), 2967.
Anthony, M. A., Bender, S. F., & van der Heijden, M. G. (2023). Enumerating soil biodiversity. Proceedings of the National Academy of Sciences, 120(33), e2304663120.
Basch G., Friedrich T., Kassam A., Gonzalez-Sanchez E. (2015). Conservation Agriculture in Europe. In: Conservation Agriculture (M. Farooq & K. H. M. Siddique, Éds.), p. 357‑389, Springer International Publishing. https://doi.org/10.1007/978-3-319-11620-4_15
Beaumelle, L., Tison, L., Eisenhauer, N., Hines, J., Maliadi, S., Pelosi, C., Philips, H.R.P. (2023). Pesticide effects on soil fauna communities—A meta‐analysis. Journal of Applied Ecology 60, 1239–1253.
Bender, S. F., Wag, C., & van der Heijden, M. G. A. (2016). An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends in Ecology & Evolution, 31
(6), 440–452. doi.org/10.1016/j.tree.2016.02.016
Berendsen, R. L., Pieterse, C. M. J., & Bakker, P. A. H. M. (2012). The rhizosphere microbiome and plant health. Trends in Plant Science, 17(8), 478–486. http://doi.org/10.1016/j.tplants.2012.04.001
Bertrand, I., Viaud, V., Daufresne, T., Pellerin, S., & Recous, S. (2019). Stoichiometry constraints challenge the potential of agroecological practices for the soil C storage. A review. Agronomy for Sustainable Development, 39, 1-16.
Bonanomi, G., Lorito, M., Vinale, F., & Woo, S. L. (2018). Organic amendments, beneficial microbes, and soil microbiota: toward a unified framework for disease suppression. Annual Review of Phytopathology, 56(1), 1-20.
Bronick, C. J. & Lal, R. (2005). Soil structure and management: a review. Geoderma 124, 3–22.
Bucaille, F. (2023). Revitaliser les sols-2e éd. : Diagnostic, fertilisation, nutriprotection. Dunod.
Bullock, J. M., Fuentes‐Montemayor, E., McCarthy, B., Park, K., Hails, R. S., Woodcock, B. A., ... & Harris, J. (2022). Future restoration should enhance ecological complexity and emergent properties at multiple scales. Ecography, 2022(4).
Chabert, A., & Sarthou, J. P. (2020). Conservation agriculture as a promising trade-off between conventional and organic agriculture in bundling ecosystem services. Agriculture, Ecosystems & Environment, 292, 106815.
Chen, H., Dai, Z., Veach, A. M., Zheng, J., Xu, J., & Schadt, C. W. (2020). Global meta-analyses show that conservation tillage practices promote soil fungal and bacterial biomass. Agriculture, Ecosystems & Environment, 293, 106841.
Christel, A., Ranjard, L., Blanchart, E., Maron, P. A. (2024). Un travail du sol ponctuel en agriculture de conservation des sols est délétère pour la vie du sol. In: L’agriculture de conservation des sols - Vrai ou faux 1. S. Cordeau, P.-A. Maron, J.-P. Sarthou, B. Chauvel, (Eds.), Editions Quae, p. 365‑369.
Cordeau S., Maron P.-A., Sarthou J.-P. et Chauvel B. (coord.). L’agriculture de conservation des sols. Versailles : Éditions Quæ (2024). 420 p. (Collection Savoir-faire). ISBN 978-2-7592-3566-7.
Crowther, L. I., Wilson, K., Wilby, A. (2023). The impact of field margins on biological pest control: a meta-analysis. BioControl, 68 (4), 387‑396.
Cousin, I. (coord.), Desrousseaux, M (coord.), Angers, D., Augusto L., …. & Leenhardt, S. (coord.) (2024). Préserver la qualité des sols : vers un référentiel d’indicateurs. Rapport d’étude, INRAE (France). www.inrae.fr/sites/default/files/pdf/IndiQuaSols_R%C3%A9sum%C3%A9%20fr%20Internet.pdf
Dainese, M., Martin, E. A., Aizen, M. A., Albrecht, M., Bartomeus, I., Bommarco, R., ... & Steffan-Dewenter, I. (2019). A global synthesis reveals biodiversity-mediated benefits for crop production. Science advances, 5(10), eaax0121.
Datnoff, L. E., Elmer, W. H. & Huber, D. M. (2007). Mineral Nutrition and Plant Disease. APS Press, St Paul, Minnesota, USA. 278p.
De Carné-Carnavalet C. (2011). Agriculture biologique : une approche scientifique. Versailles : Éditions France Agricole, collection Agriproduction. 472 p. ISBN 978-2-85557-211-6.
de Graaff M.-A., Hornslein N., Throop H. L., Kardol P., van Diepen L. T. A. (2019). Effects of agricultural intensification on soil biodiversity and implications for ecosystem functioning: A meta-analysis. Advances in Agronomy Vol. 155, p. 1‑44, Elsevier. https://doi.org/10.1016/bs.agron.2019.01.001
Deguine, J.-P., Aubertot, J.-N., Bellon, S., Côte, F.-X., …& Lamichhane, J.R. (2023). Agroecological crop protection for sustainable agriculture. Advances in Agronomy 178, 1–59.
Derksen D. A., Blackshaw R. E., Boyetchko S. M. (1996). Sustainability, conservation tillage and weeds in Canada. Canadian Journal of Plant Science
, 76 (4), 651‑659. doi.org/10.4141/cjps96-115
Derrouch, D., Dessaint, F., Chauvel, B. (2024). Gestion de la flore adventice en agriculture de conservation des sols : retours d’expérience d’agriculteurs. In: L’agriculture de conservation des sols - Chapitre 10. S. Cordeau, P.-A. Maron, J.-P. Sarthou, B. Chauvel, (Eds.), Editions Quae, p. 215‑227.
Devi, R., Kaur, T., Kour, D., Yadav, A., Yadav, A.N., Suman, A., Ahluwalia, A.S., Saxena, A.K. (2022). Minerals solubilizing and mobilizing microbiomes: A sustainable approach for managing minerals’ deficiency in agricultural soil. Journal of Applied Microbiology133, 1245–1272.
Dini-Andreote, F., Stegen, J. C., van Elsas, J. D. & Salles, J. F. (2015). Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. U.S.A. 112, E1326–E1332.
Domeignoz-Horta, L. A., Cappelli, S. L., Shrestha, R., Gerin, S., Lohila, A. K., Heinonsalo, J., ... & Laine, A. L. (2024). Plant diversity drives positive microbial associations in the rhizosphere enhancing carbon use efficiency in agricultural soils. Nature Communications, 15(1), 8065.
Duru M., Sarthou, J. P., Therond, O. (2022). L’agriculture régénératrice : summum de l’agroécologie ou greenwashing ? Cahiers Agricultures, 31, 10p.
Duru, M., Therond, O., Martin, G., Martin-Clouaire, R., Magne, M. A., Justes, E., ... & Sarthou, J. P. (2015). How to implement biodiversity-based agriculture to enhance ecosystem services: a review. Agronomy for sustainable development, 35, 1259-1281.
EEA, 2002. An inventory of biodiversity indicators in Europe. Technical report, n° 92, 2 pp.
Emmett, B., Büchi, L., Smith, B., Soleiman, J., Thompson, W., & Dodsworth, J. (2025). Regenerative agriculture in the UK: an ecological perspective.
EU (2018). Regulation (EU) 2018/848 of the European Parliament and of the Council of 30 May 2018 on organic production and labelling of organic products and repealing Council Regulation (EC) No 834/2007. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32018R0848
Fanin, N., Kardol, P., Farrell, M., Nilsson, M-C., Gundale, M.J., Wardie, D.A. (2019). The ratio of Gram-positive to Gram-negative bacterial PLFA markers as an indicator of carbon availability in organic soils. Soil Biology and Biochemistry 128, 111–114.
Fierer, N., Jackson, R. B. (2006). The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences, 103 (3), 626‑631. https://doi.org/10.1073/pnas.0507535103
Giller, K.en, Hijbeek, R., Anderson, J.A., Sumberg, J. (2021). Regenerative agriculture: an agronomic perspective. Outlook on Agriculture. 50 (1), 13-25. DOI: 10.1177/0030727021998063
Gliessman, S. R., Méndez V. E, Izzo Victor M., Engles, W. (2022). Agroecology: Leading the Transformation to a Just and Sustainable Food System. CRC Press, 470 pages.
Guyonnet, JP., Guillemet, M., Dubost, A., et al. (2018). Plant nutrient resource use strategies shape active rhizosphere microbiota through root exudation. Front Plant Sci 871:1–13. https:// doi. org/ 10. 3389/ fpls. 2018. 01662
Holmes, K. D., Blubaugh, C. K. (2023). A Guide to 23 Global Syntheses of Plant Diversity Effects: Unpacking Consensus and Incongruence across Trophic Levels. The Quarterly Review of Biology, 98 (3), 121‑148. https://doi.org/10.1086/726687
Hossain, M. B. (2021). Glomalin and Contribution of Glomalin to Carbon Sequestration in Soil: A Review. Food Science and Technology, 9 (1), 191‑196. htps://doi.org/10.24925/turjaf.v9i1.191-196.3803
Howe, J. A., McDonald, M.D., Burke, J., Robertson, I., Coker, H., Gentry, T., Lewis, K.L. (2024). Influence of fertilizer and manure inputs on soil health: A review. Soil Security 16, 100155.
Husson, O. (2013). Redox potential (Eh) and pH as drivers of soil/plant/microorganism systems: a transdisciplinary overview pointing to integrative opportunities for agronomy. Plant and Soil, 362 (1-2): 389-417. https://doi.org/10.1007/s11104-012-1429-7
Husson, O. (2023). How pH and Eh Influence Soil Nutrient Dynamics with Microbial Mediation. In: Biological approaches to regenerative soil systems. Uphoff, N. and Thies, J. (Eds). CRC Press, New York. Chapter 21. ISBN: 9780367554712
Husson, O., Husson, B., Brunet, A., Babre, D., Alary, K., Sarthou, J.-P., Charpentier, H., Durand, M., Benada, J., Henry, M. (2016). Practical improvements in soil redox potential (Eh) measurement for characterisation of soil properties. Application for comparison of conventional and conservation agriculture cropping systems. Analytica Chimica Acta
906, 98–109. doi.org/10.1016/j.aca.2015.11.052
Husson, O., Sarthou, J.-P., Bousset, L., Ratnadass, A., Schmidt, H.-P., Kempf, J., Husson, B., Tingry, S., Aubertot, J.-N., Deguine, J.-P., Goebel, F.-R., Lamichhane, J.R. (2021). Soil and plant health in relation to dynamic sustainment of Eh and pH homeostasis: A review. Plant Soil 466, 391–447. https://doi.org/10.1007/s11104-021-05047-z
Husson, O., Brunet, A., Babre, D., Charpentier, H., Durand, M., Sarthou, J.-P. (2018). Conservation Agriculture systems alter the electrical characteristics (Eh, pH and EC) of four soil types in France. Soil and Tillage Research176, 57–68.
IFOAM. (2005). Principles of Organic Agriculture. International Federation of Organic Agriculture Movements, Bonn, Germany. https://www.ifoam.bio/sites/default/files/2020-01/poa_english_web.pdf
Islam Z. F., Greening C., Hu H. (2023). Microbial hydrogen cycling in agricultural systems – plant beneficial or detrimental? Microbial Biotechnology, 16 (8), 1623‑1628. https://doi.org/10.1111/1751-7915.14300
Jacoby, R., Peukert, M., Succurro, A., Koprivova, A., Kopriva, S. (2017). The Role of Soil Microorganisms in Plant Mineral Nutrition—Current Knowledge and Future Directions. Frontiers in Plant Science, 8(September), 1–19. http://doi.org/10.3389/fpls.2017.01617
Jasrotia, P., Kumari, P., Malik, K., Kashyap, P.L., Kumar, S., Bhardwaj, A.K., Singh, G.P. (2023). Conservation agriculture based crop management practices impact diversity and population dynamics of the insect-pests and their natural enemies in agroecosystems. Frontiers in Sustainable Food Systems 7, 1173048
Jayaraman, S., Naorem, A. K., Lal, R., Dalal, R. C., Sinha, N. K., Patra, A. K., Chaudhari, S. K. (2021). Disease-Suppressive Soils—Beyond Food Production: a Critical Review. Journal of Soil Science and Plant Nutrition, 21 (2), 1437‑1465.
Jeyaseelan, A. (2024). A review of the impact of herbicides and insecticides on the microbial communities. Environmental Research. 245, 118020.
Johannes, A., Matter, A., Schulin, R., Weisskopf, P., Baveye, P. C., Boivin, P. (2017). Optimal organic carbon values for soil structure quality of arable soils. Does clay content matter? Geoderma, 302(April), 14–21. http://doi.org/10.1016/j.geoderma.2017.04.021
Johnston, A. E., Poulton, P. R. (2018). The importance of long‐term experiments in agriculture: their management to ensure continued crop production and soil fertility; the Rothamsted experience. European Journal of Soil Science
, 69 (1), 113‑125. doi.org/10.1111/ejss.12521.
Karp, D. S., Chaplin-Kramer, R., Meehan, T. D., Martin, E. A., DeClerck, F., Grab, H., ... & Wickens, J. B. (2018). Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proceedings of the National Academy of Sciences, 115(33), E7863-E7870.
Kesavan, P. C., Malarvannan, S. (2010). Green to evergreen revolution: ecological and evolutionary perspectives in pest management. Current Science, 99 (7), 908‑914.
Kesse-Guyot, E., Baudry, J., Assmann, K., Galan, P., Hercberg, S., Lairon, D. (2017). Prospective Association between Consumption Frequency of Organic Food and Body Weight Change, Risk of Overweight or Obesity: Results from the NutriNet-Santé Study. British Journal of Nutrition 117 (02): 325–34. doi:10.1017/S0007114517000058.
Khangura, R., Ferris, D., Wag, C., Bowyer, J. (2023). Regenerative Agriculture — A Literature Review on the Practices and Mechanisms Used to Improve Soil Health. Sustainability, 1–43.
Kirkby, C. A., Richardson, A. E., Wade, L. J., Batten, G. D., Blanchard, C., Kirkegaard, J. A. (2013). Carbon-nutrient stoichiometry to increase soil carbon sequestration. Soil Biology and Biochemistry, 60, 77‑86. https://doi.org/10.1016/j.soilbio.2013.01.011
Kirkby, C. A., Richardson, A. E., Wade, L. J., Conyers, M., & Kirkegaard, J. A. (2016). Inorganic nutrients increase humification efficiency and C-sequestration in an annually cropped soil. PLoS One, 11(5), e0153698.
Kirkby, C. A., Richardson, A. E., Wade, L. J., Passioura, J. B., Batten, G. D., Blanchard, C., Kirkegaard, J. A. (2014). Nutrient availability limits carbon sequestration in arable soils. Soil Biology and Biochemistry, 68, 402‑409. https://doi.org/10.1016/j.soilbio.2013.09.032
Kirschbaum, M. U. F., Moinet, G. Y. K., Hedley, C. B., Beare, M. H., McNally, S. R. (2020). A conceptual model of carbon stabilisation based on patterns observed in different soils. Soil Biology and Biochemistry, 107683. https://doi.org/10.1016/j.soilbio.2019.107683
Kutcher, H. R., Johnston, A. M., Bailey, K. L., Malhi, S. S. (2011). Managing crop losses from plant diseases with foliar fungicides, rotation and tillage on a Black Chernozem in Saskatchewan, Canada. Field Crops Research, 124, 205‑212.
Lavelle, P., Spain, A., Fonte, S., Bedano, J.C., Blanchart, E., Galindo, V., Grimaldi, M., Jimenez, J.J., Velasquez, E., Zangerié, A. (2020) Soil aggregation, ecosystem engineers and the C cycle. Acta Oecologica105, 103561.
La Scala, N. J, Lopes, A., Spokas, K ., Bolonhezi, D., Archer, D.W., Reicosky, D.C. (2008). Short-term temporal changes of soil carbon losses after tillage described by a first-order decay model. Soil and Tillage Research99, 108–118.
Lebeis, S. L., Paredes, S. H., Lundberg, D. S. (2015). Break eld N, Gehring J, McDonald M, Malfatti S, Del Rio TG, Jones CD, Tringe SG: Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science, 349, 860-864.
Lefèvre, C. (2024). Agroecological drivers connecting soil management practices, soil health, plant health and the nutritional and sanitary qualities of products: the case of soft winter wheat to illustrate the “One Health” approach. PhD thesis, Université de Toulouse, 189 pp.
Lehmann, J., & Kleber, M. (2015). The contentious nature of soil organic matter. Nature, 528(7580), 60-68.
Lescuyer T, Duru M, Wallet F (2025). Agriculture régénératrice et nouvelles filières : quel potentiel de territorialisation ? (soumis)
Letourneau, D. K., Armbrecht, I., Rivera, B. S., Lerma, J. M., Carmona, E. J., Daza, M. C., ... & Trujillo, A. R. (2011). Does plant diversity benefit agroecosystems? A synthetic review. Ecological applications, 21(1), 9-21.
Liang, Y., Cao, D., Ma, Z., Wu, R., Zhang, H., Fang, Y., ... & Zhu, Z. (2024). Stoichiometry regulates rice straw-induced priming effect: The microbial life strategies. Soil Biology and Biochemistry, 196, 109514
Lin D., McCulley R. L., Nelson J. A., Jacobsen K. L., Zhang D. (2020). Time in pasture rotation alters soil microbial community composition and function and increases carbon sequestration potential in a temperate agroecosystem. Science of The Total Environment, 698, 134233. https://doi.org/10.1016/j.scitotenv.2019.134233
Löbmann, M. T., Vetukuri, R. R., de Zinger, L., Alsanius, B. W., Grenville-Briggs, L. J., Walter, A. J. (2016). The occurrence of pathogen suppressive soils in Sweden in relation to soil biota, soil properties, and farming practices. Applied Soil Ecology, 107, 57‑65. https://doi.org/10.1016/j.apsoil.2016.05.011
Lupwayi, N. Z., Turkington, K. T., Tidemann, B. D., Kubota, H. & Polo, R. O. (2024). Crop rotation and fungicides impact microbial biomass, diversity and enzyme activities in the wheat rhizosphere. Frontiers in Agronomy 6, 1455448.
Mambo, T., Lhermie, G. (2024). “The Futures for Regenerative Agriculture: Insights from the Organic Movement and the Tussle with Industrial Agriculture.” Frontiers in Sustainable Food Systems 8 (December): 1–10. doi:10.3389/fsufs.2024.1455024.
McLaughlin, A., Mineau, P. (1995). The impact of agricultural practices on biodiversity. Agriculture, Ecosystems & Environment 55, 201–212.
Meghvansi, M.K., Singh, L., Srivastava, R.B., Varma, A. (2011). Assessing the Role of Earthworms in Biocontrol of Soil-Borne Plant Fungal Diseases. In: Karaca, A. (eds) Biology of Earthworms. Soil Biology, vol 24. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-14636-7_11
Mehra, P., Baker, J., Sojka, R.F., Bolan, N., Desbiolles, J., Kirkham, M.B., Ross, C., Gupta, R. (2018). A Review of Tillage Practices and Their Potential to Impact the Soil Carbon Dynamics. Advances in Agronomy 150, 185–230.
Modak, K., Ghost, A., Bhattacharyya, R., Biswas, D.K., Das, T.K., Das, S., Singh, C. (2019). Response of oxidative stability of aggregate-associated soil organic carbon and deep soil carbon sequestration to zero-tillage in subtropical India. Soil & Tillage Research195, 104370.
Newbold, T., Hudson, L. N., Hill, S. L., Contu, S., Lysenko, I., Senior, R. A., ... & Purvis, A. (2015). Global effects of land use on local terrestrial biodiversity. Nature, 520(7545), 45-50.
Noreen, S., Fatima, Z., Ahmad, S., Athar, H.-R. & Ashraf, M. (2018) Foliar Application of Micronutrients in Mitigating Abiotic Stress in Crop Plants. in Plant nutrients and abiotic stress tolerance 95–116. Springer Nature, Singapore.
Panagos, P., Borrelli, P., Jones, A., & Robinson, D. A. (2024). A 1 billion euro mission: A Soil Deal for Europe. European Journal of Soil Science, 75(1), e13466.
Paolini, A., Bhalla, I., & Loring, P. (2024). “Doing more good”: Exploring the multidisciplinary landscape of regeneration as a boundary object for paradigm change. Journal of Agriculture, Food Systems, and Community Development, 13(4), 1-16.
Peng, J., Liu,H., Hu, Y., Sun, Y., Liu, Q., Li,J., Dong, Y. (2022). Shift in soil bacterial communities from K ‐ to r ‐strategists facilitates adaptation to grassland degradation. Land Degrad Dev 33, 2076–2091.
Pieterse, C. M., Zamioudis, C., Berendsen, R. L., Weller, D. M., Van Wees, S. C., Bakker, P. A. (2014). Induced systemic resistance by beneficial microbes. Annual review of phytopathology, 52, 347-375.
Poeplau, C., Don, A. (2023). A simple soil organic carbon level metric beyond the organic carbon‐to‐clay ratio. Soil Use and Management, 39(3), 1057-1067.
Power, A. G. (2010). Ecosystem services and agriculture: tradeoffs and synergies. Philosophical transactions of the royal society B: biological sciences, 365(1554), 2959-2971.
Prăvălie, R., Borrelli, P., Panagos, P., Ballabio, C., Lugato, E., Chappell, A., …. & Birsan, M.-V. (2024). A Unifying Modelling of Multiple Land Degradation Pathways in Europe. Nature Communications
15:3862. doi.org/10.1038/s41467-024-48252-x
Rabbi, S. M. F., Minasny, B., McBratney, A. B., Young, I. M. (2020). Microbial processing of organic matter drives stability and pore geometry of soil aggregates. Geoderma, 360, 114033.
Rabot, E., Saby, N. P., Martin, M. P., Barré, P., Chenu, C., Cousin, I., ... & Bispo, A. (2024). Relevance of the organic carbon to clay ratio as a national soil health indicator. Geoderma, 443, 116829
Rasmusson, A. G., Escobar, M. A., Hao, M., Podgórska, A. & Szal, B. (2020). Mitochondrial NAD(P)H oxidation pathways and nitrate/ammonium redox balancing in plants. Mitochondrion53, 158–165.
Rehberger, E., West, P. C., Spillane, C., & McKeown, P. C. (2023). What climate and environmental benefits of regenerative agriculture practices? an evidence review. Environmental Research Communications, 5(5), 052001.
Rigal, S., Dakos, V., Alonso, H., Auniņš, A., Benkő, Z., Brotons, L., ... & Devictor, V. (2023). Farmland practices are driving bird population decline across Europe. Proceedings of the National Academy of Sciences, 120(21), e2216573120.
Rowley, M. C., Grand S., Verrecchia É. P. (2018). Calcium-mediated stabilisation of soil organic carbon. Biogeochemistry, 137 (1‑2), 27‑49.
Sarthou, J. P., Boulakia, S., Singla, S., Cordeau, S., Tivet, F. (2024a). Approche système de la mise en œuvre des piliers de l’agriculture de conservation des sols. In: L’agriculture de conservation des sols - Chapitre 2. S. Cordeau, P.-A. Maron, J.-P. Sarthou, B. Chauvel (Eds.), Editions Quae : 51‑77.
Sarthou, J.-P., Gardarin, A., Waligora, C., Blanchart, E. (2024b). Gestion des ravageurs des cultures en agriculture de conservation des sols. In: L’agriculture de conservation des sols - Chapitre 9. S. Cordeau, P.-A. Maron, J.-P. Sarthou, B. Chauvel (Eds.), Editions Quae : 189‑214.
Sarthou, J.-P., Sester, M., Lamichhane, J. R., Aubertot, J.-N., Sarthou, V., Suffert, F. (2024c). Effets des pratiques d’agriculture de conservation des sols sur les maladies cryptogamiques. In: L’agriculture de conservation des sols - Chapitre 8. S. Cordeau, P.-A. Maron, J.-P. Sarthou, B. Chauvel (Eds.), Editions Quae : 173‑187.
Sauzet, O., Johannes, A., Deluz, C., Dupla, X., Matter, A. Baveye P.C., Boivin, P. (2024). The organic carbon‐to‐clay ratio as an indicator of soil structure vulnerability, a metric focused on the condition of soil structure. Soil Use and Management 40, e13060.
Sias, C., Wolters, B. R., Reiter, M. S. & Flessner, M. L. (2021). Cover crops as a weed seed bank management tool: A soil down review. Italian Journal of Agronomy 16, 1852.
Seibold, S., Gossner, M. M., Simons, N. K., Blüthgen, N., Müller, J., Ambarlı, D., ... & Weisser, W. W. (2019). Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature, 574(7780), 671-674.
Sela, S., Dobermann, A., Cerri, C. E., Svoray, T., van-Es, H., Amsili, J., ... & Katz, S. (2024). Towards a unified approach to prioritization of regenerative agricultural practices across cropping systems. npj Sustainable Agriculture, 2(1), 24.
Shackelford, G., Steward, P. R., Benton, T. G., Kunin, W. E., Potts, S. G., Biesmeijer, J. C., & Sait, S. M. (2013). Comparison of pollinators and natural enemies: a meta‐analysis of landscape and local effects on abundance and richness in crops. Biological Reviews, 88(4), 1002-1021.
Sher, A., Li, H., Ullah, A., Hamid, Y., Nasir, B., Zhang, J. (2024). Importance of regenerative agriculture: climate, soil health, biodiversity and its socioecological impact. Discover Sustainability5, 462.
Smith, R. G., Gross, K. L., Robertson, G. P. (2008). Effects of Crop Diversity on Agroecosystem Function: Crop Yield Response. Ecosystems, 11, 355‑366.
Soong, J. L., Fuchslueger, L., Marañon-Jimenez, S., Torn, M. S., Janssens, I. A., Penuelas, J., & Richter, A. (2020). Microbial carbon limitation: The need for integrating microorganisms into our understanding of ecosystem carbon cycling. Global Change Biology, 26(4), 1953–1961. http://doi.org/10.1111/gcb.14962
Staton, T., Walters, R. J., Smith, J., & Girling, R. D. (2019). Evaluating the effects of integrating trees into temperate arable systems on pest control and pollination. Agricultural Systems, 176, 102676.
Steenwyk, P., Heun, M. K., Brockway, P., Sousa, T., & Henriques, S. (2022). The contributions of muscle and machine work to land and labor productivity in world agriculture since 1800. Biophysical Economics and Sustainability, 7(2), 2.
Stuart, E. K., Castañeda-Gómez, L., Buss, W., Powell, J. R. & Carrillo, Y. (2024). Non-mycorrhizal root-associated fungi increase soil C stocks and stability via diverse mechanisms. Biogeosciences 21, 1037–1059.
Tamburini, G., Bommarco, R., Wanger, T. C., Kremen, C., van der Heijden, M. G. A., Liebman, M., Hallin, S. (2020). Agricultural diversification promotes biodiversity and multiple ecosystem services without compromising yield. Science Advances, 2020; 6: eaba1715.
Tao F., Huang Y., Hungate B. A., Manzoni S., Frey S. D., Schmidt M. W. I., Reichstein M., Carvalhais N., Ciais P., Jiang L., Lehmann J., Wang Y.-P., Houlton B. Z., Ahrens B., Mishra U., Hugelius G., Hocking T. D., Lu X., Shi Z., … Luo Y. (2023). Microbial carbon use efficiency promotes global soil carbon storage. Nature, 618 (7967), 981‑985. https://doi.org/10.1038/s41586-023-06042-3
Therond, O., Duru, M. (2019). Agriculture et biodiversité : les services écosystémiques, une voie de réconciliation? Innovations agronomiques, 75, 29-47.
Therond, O., Duru, M., Roger-Estrade, J., Richard, G. (2017). A new analytical framework of farming system and agriculture model diversities. A review. Agronomy for sustainable development, 37, 1-24.
Tittonell, P., Mujtar, V. El, Felix, G., Kebede, Y., Laborda, L., & Soto, R. L. (2022). Regenerative agriculture — agroecology without politics ? Sustainable Food Systems, 6, 844261.
Tyc, O., van den Berg, M., Gerards, S., van Veen, J.A., Raaijmakers, J.M., De Boer, W., Garbeva, P. (2014). Impact of interspecific interactions on antimicrobial activity among soil bacteria. Frontiers in Microbiology5, article 567.
Van Elsas, J. D., Chiurazzi, M., Mallon, C. A., Elhottovā, D., Krištůfek, V., Salles, J. F. (2012). Microbial diversity determines the invasion of soil by a bacterial pathogen. Proceedings of the National Academy of Sciences, 109 (4), 1159‑1164.
Van Groenigen, J. W., Lubbers, I. M., Vos, H. M., Brown, G. G., De Deyn, G. B., & Van Groenigen, K. J. (2014). Earthworms increase plant production: a meta-analysis. Scientific reports, 4(1), 6365.
Vialatte, A., Martinet, V., Tibi, A., Alignier, A., Angeon, V., et al., (2023). Protéger les cultures en augmentant la diversité végétale des espaces agricoles. Rapport scientifique de l’Expertise scientifique collective. INRAE. 2023, 954 p. ⟨hal-04127709v4⟩
Virk, A. L., Shakoor, A., Abdullah, A., Chang, S. X. & Cai, Y. (2024). Pesticide effects on crop physiology, production and soil biological functions. Advances in Agronomy187, 171–212
Wang, G., Li, X., Xi, X., & Cong, W. F. (2022). Crop diversification reinforces soil microbiome functions and soil health. Plant and Soil, 476(1), 375-383.
Wang, B. (2021). Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biology and Biochemistry.
Wang, X., Chi, Y. & Song, S. (2024). Important soil microbiota’s effects on plants and soils: a comprehensive 30-year systematic literature review. Frontiers in Microbiology15, 1347745.
Wortmann C. S., Dang Y. P. (2020). Strategic Tillage for the Improvement of No-Till Farming Systems. In: No-till Farming Systems for Sustainable Agriculture (Y. P. Dang, R. C. Dalal, & N. W. Menzies, Éds.), p. 155‑171, Springer International Publishing. doi.org/10.1007/978-3-030-46409-7_10
Xu Z., Tsang D. C. W. (2024). Mineral-mediated stability of organic carbon in soil and relevant interaction mechanisms. Eco-Environment & Health, 3 (1), 59‑76. https://doi.org/10.1016/j.eehl.2023.12.003
Xu, Q., Li, L., Guo, J., Liu, M., Guo, S., Kuzyakov, Y., Ling, N., Shen, Q., Martiny, J.B.H., Hungate, B.A. (2024). Active microbial population dynamics and life strategies drive the enhanced carbon use efficiency in high-organic matter soils. Microbial ecology15, 1–20.
Yu G.-H., Kuzyakov Y. (2021). Fenton chemistry and reactive oxygen species in soil: Abiotic mechanisms of biotic processes, controls and consequences for carbon and nutrient cycling. Earth-Science Reviews, 95 (14), 106408. https://doi.org/10.1016/j.earscirev.2021.103525
Zahm F., Vilain L., Barbier J.-M., Bellon S., Doreau M., Gasselin P., Viaux P. et Parguel M. (2020). La méthode IDEA – Indicateurs de durabilité des exploitations agricoles. Guide d’utilisation (version 4). Educagri éditions, 252 pp.
Zhang, Y. Ye, C., Su, Y. Peng, W., Lu, R. Liu, Y.Huang, H., He, X., Yang, M. Zhu, S. (2022). Soil acidification caused by excessive application of nitrogen fertilizer aggravates soil-borne diseases: Evidence from literature review and field trials. Agriculture, Ecosystems and Environment 340, 108176.
Zhu, Z., Fang, Y., Liang, Y., Li, Y., Liu, S., Li, Y., ... & Ge, T. (2022). Stoichiometric regulation of priming effects and soil carbon balance by microbial life strategies. Soil Biology and Biochemistry, 169, 108669.
Zulfiqar, F., Russell, G., Hancock, J. T. (2021). Molecular hydrogen in agriculture. Planta, 254 (3), 56. https://doi.org/10.1007/s00425-021-03706-0

Les articles sont publiés sous la licence Creative Commons (CC BY-NC-ND 2.0)
Pour la citation et la reproduction de cet article, mentionner obligatoirement le titre de l'article, le nom de tous les auteurs, la mention de sa publication dans la revue AES et de son URL, la date de publication.